Поиск по сайту
- Вы здесь:
-
Главная

-
book
-
Группы крови человека
- Система Kell
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...

- Категория: Система Kell
Частота распределения антигена К у разных народов имеет определенную закономерность. В России по мере продвижения с Запада на Восток, из Европы в Азию, по оси Москва-Токио частота фактора К постепенно убывает: русские (Смоленск) - 8,85 %; русские (Нижегородская область) - 8,7 %; русские (Москва) - 8,1 %, русские (Сургут) - 6,21 %; русские (Первоуральск) - 6,0 %; коми - 6,04%, манси - 1,06%, ханты поселковые - 4,1 %, ханты лесные -0,32 %, буряты - 0,22 %, монголы - 0,4 %, китайцы, корейцы и японцы - 0 %.
В распределении групповых антигенов эритроцитов среди разных народов можно выделить монголоидную триаду, характеризующую представителей монголоидной расы: высокая частота группы 0(1) или В(Ш), высокая частота антигена D, низкая частота антигена К. Распределение антигена К среди населения России подтверждает это положение. В Европейской части России, где подавляющее большинство жителей относятся к европеоидам, частота антигена К наиболее высокая, в Азиатской части России, где преимущественно проживают монголоиды, она приближается к 0.
Антиген Кра (Penney) у европеоидов, в том числе у русских, встречается с частотой около 2 %, антиген Kpb (Rautenberg) - в 100 % случаев. У негров в 20 раз чаще, чем у европейцев встречается антиген Jsa (Sutter) - 19,5 % и < 1 % соответственно. Антиген Ula (Karhula) распространен практически только среди финнов. Частота его в этой популяции составляет 2,6 % [169], у шведов - 0,2 % [169]. Среди обследованных 5 тыс. англичан антиген 1Ла не найден [169], среди 8 тыс. японцев частота этого антигена составила 0,46 % [300].
Частота антигена К у разных народов
|
Популяция |
Объем выборки |
Частота фенотипа, % |
|
|
|
к+ Г |
К- |
Источник |
||
|
Абхазы |
116 |
11,2 |
88,8 |
Саламатина Н.В. и соавт. [62] |
|
Аджарцы |
485 |
0,02 |
99,98 |
Нагервадзе М.А. и соавт. [50, 296] |
|
Азербайджанцы |
1 690 |
5,3 |
94,6 |
Таги-заде Р.К. и соавт. [69-71] |
|
Алеуты |
20 40 |
0,0 2,5 |
100 97,5 |
Рынков Ю.Г., Шереметьева В.А. [60,61] |
|
Алтайцы северные |
56 508 179 |
0,0 7,3 14 |
100 92,7 86,0 |
Сукерник Р.И. и соавт. [66-68] |
|
Армяне |
1 152 395 |
8,56 8,6 |
91,44 91,4 |
Нерсисян В.М. и соавт. [51-53] |
|
Башкиры |
93 64 |
0,0 7,8 |
100 92,2 |
Рафиков Х.С. и соавт. [59] |
|
Белорусы |
74 93 |
8,1 14 |
91,9 86 |
Лопатенок А.А., Будяков О.С. [42] Микулич А.И. и соавт. [45] |
|
Буряты |
99 4 222 |
0,0 0,22 |
100 99,78 |
Архив КА МГУ № 23 (цит. по Ю.Г. Рычкову и соавт. [13]) Зеленцова В.Ф., Бурлаева Э.М. [36,37] |
|
Грузины |
40 659 |
5,0 5,8 |
95 94,2 |
Kherumian и соавт., (цит. по Ю.Г. Рычкову и соавт. [13]) Вербицкий М.Ш. и соавт. [11] |
|
Дагестанцы |
45 38 |
0,0 7,9 |
100 92,1 |
Гаджиев А.Г. [12] |
|
Ительмены |
18 |
10 и выше |
Меньше 100 |
Архив КА МГУ № 27 (цит. по Ю.Г. Рычкову и соавт. [13]) |
|
Казахи |
106 28 |
18,9 28,6 |
81,1 71,4 |
Архив ЛГЧ ИОГен, № 136 (цит. по Ю.Г. Рычкову и соавт. [13]) |
|
Калмыки |
А-79 |
Ш 1 |
98,7 |
Politzer, 1962 (цит. по Ю.Г. Рычкову и соавт. [131) |
|
Киргизы |
454 762 |
3,96 3,14 |
96,04 96,85 |
Буренкова Л.В. [10] Килина Г.А., Турсунбаев М.С. [39] |
|
Коми |
105 123 92 4 885 |
0,0 7,3 25 6,04 |
100 92,7 75 93,96 |
Эрикссон А., Франте Р. [83] Мороков В.А. [48,49] |
|
Коми-пермяки |
107 |
6,5 |
93,5 |
Архив ЛГЧ ИОГен, № 102 (цит. по Ю.Г. Рычкову и соавт. [13]) |
|
Коряки |
37 16-208 |
0,0 до 10 |
100 меньше 100 |
Архив КА МГУ № 24 (цит. по Ю.Г. Рычкову и соавт. [13]) Шереметьева В.А., Горшков В.А. [81] |
|
Манси |
59 126 |
0,0 32 |
100 96,8 |
Архив ЛГЧ ИОГен, № 137 (цит. по Ю.Г. Рычкову и соавт. [13]) Давыдова Г.М. [16] |
|
К+ |
к- |
Источник |
||
|
Марийцы |
103 117 |
2 4,3 |
98 95,7 |
Эрикссон А. и соавт. [82] |
|
Нганасаны |
195 235 |
0,0 0,0 |
100 100 |
Сукерник РИ. и соавт. [66] |
|
Нивхи |
10-21 |
до 30 |
70-85,7 |
Бахолдина В.Ю. [4] |
|
Русские, Дзержинск |
8 747 |
8,7 |
91 |
Червяков В.И. [79] |
|
Русские, Москва |
1 017 |
7,8 |
92 |
Умнова М.А. [74- 77] |
|
Русские, Москва |
5 617 |
8 |
92 |
Башлай А.Г. [5] |
|
Русские Москва |
1258 |
8,0 |
92 |
Пискунова Т.М. [57] |
|
Русские, Смоленск |
9 997 |
8,8 |
91 |
Михайлова Н.М. [46,47] |
|
Русские, Сургут |
9 376 |
6,2 |
93,7 |
. Меркулова Н.Н. [43,44] |
|
Русские, Первоуральск |
3 577 |
6,0 |
94 |
Скудицкий А.Е. [63] |
|
Русские |
102 5 617 100 537 |
4,9 8,1 8,7 |
95.1 91,9 91,3 |
Бронникова М.А., Барсегянц Л.О. [9] Башлай А.Г. [6] Червяков В.И. [79] |
|
Татары |
50 |
2 |
96 |
Лопатенок А.А., Будяков О.С. [42] |
|
Узбеки |
50 126 |
4,0 Ш 34,9 |
96,0 65,1 |
Лопатенок А.А., Будяков О.С. [42м jjj Архив ЛГЧ ИОГен, №138 (цит. по Ю.Г. Рычкову и соавт. [13]) | |
|
Украинцы |
330 392 |
4,6 5,8 |
95,4 94,2 |
Лопатенок А.А., Будяков О.С. [42] 1 Chown, Lewis (цит. по Ю.Г Рычкову и соавт. [13])| |
|
Хакасы |
137 |
9,5 |
90,5 |
Перевозчиков И.В. и соавт. [56] |
|
Хакасы всего, в том числе: |
429 |
4,0 |
95 |
Абдина А.С. [2] |
|
кызыльцы |
74 |
100 |
||
|
качинцы |
60 |
5,0 |
94 |
|
|
койбалы |
74 |
100 |
||
|
сагайцы |
78 |
1,3 |
98 |
|
|
бирюсинцы |
70 |
8,6 |
91 |
|
|
бельтыры |
73 |
5,3 |
94 |
|
|
Чукчи |
214 46 |
0,0 10,9 |
100 89,1 |
Сукерник Р.И. и соавт. [67] Рычков Ю.Г, Шереметьева В.А. [60] |
|
Эвенки восточные |
12-41 |
10 и выше |
меньше 100 |
Архив КА МГУ № 24,25,29,38 1 (цит. по Ю.Г. Рычкову и соавт. [13]) |
|
Эвенки западные |
214 59 |
11,2 0,0 |
88,8 100 |
Ю.Г. Рычков и соавт. [61] Архив ЛГЧ ИОГен, № 139 (цит. по Ю.Г. Рычкову и соавт. [13]) |
|
Эвены |
6 15-34 |
0,0 10 и выше |
100 меньше 100 |
| Архив КА МГУ № 24 (цит. по Ю.Г. Рычкову и соавт. [13])| |
|
Эскимосы |
102 Щ21-34 |
0,0 2,9^1,3 |
100 85,7-97,1 |
Сукерник Р.И. и соавт. [68] Рычков Ю.Г, Шереметьева В.А. [601 |
|
К+ |
к- |
|||
|
Якуты |
57 241 |
0,0 3,7 |
100 96,3 |
Архив ЛГЧ ИОГен, № 139 (цит. по Ю.Г. Рычкову и соавт. [13]) Золотарева И.М., Башлай А.Г. [38] |
|
Негры |
4 079 |
1,5 |
98,5 |
Stroup at al. [357] |
|
Англичане |
9 875 |
9,0 |
|
Race, Sanger [318] |
|
Французы |
81962 |
8,5 |
|
Garetta и соавт. [172] |
|
Финны |
5 000 |
4,1 |
|
Fnruhjelm U. и соавт. [169] |
|
Чеченцы |
2 389 |
1,6 |
98,3 |
АбаеваЛ.М. [1] |
|
Удмурты |
2 404 |
2,2 |
97,7 |
Суворов А.В. и соавт. [65] |
|
Ханты |
302 |
0,3 |
99,7 |
Хромова Е.А. [78] |
|
Монголы |
490 |
0,4 |
99,6 |
Чимиддуламын Ш. [80] |
Данные литературы, касающиеся частоты факторов Kell среди представителей одного и того же народа, нередко противоречивы даже в публикациях одних и тех же авторов. Например, у алеутов, северных алтайцев, башкир и бурят (табл. 5.6, строки 4, 5, 7, 9) антиген К в одних выборках не обнаружен, в то время как в других выборках обнаружен с частотой более 7 %. Еще большие различия (от 0 до 14 %) прослеживаются у дагестанцев, коряков, коми, чукчей, эвенков, эвенов, эскимосов и якутов (см. табл. 5.6, строки 11, 16, 18, 28-33). У белорусов (см. табл. 5.6, строка 8) в одной выборке частота антигена К составила 8,1 %, в другой - 14 %; у казахов (см. табл. 5.6, строка 13) - 18,9 и 28,6 %; у узбеков (см. табл. 5.6, строка 25) - 4 и 34,9 %.
Таблица 5.7
Частота антигенов Кра и Крь у разных народов
|
Популяция |
Объем выборки |
Частота фенотипа, % |
Источник |
||
|
Кр(а+Ь-) |
Кр(а+Ь+) |
Кр(а-Ь+) |
|||
|
Русские |
512 560 |
1,8 |
100 |
Пискунова Т.М. и соавт. [58] |
|
|
Коми |
92-123 |
1,9-3,3 |
96,7-98,1 |
Эрикссон А., Франте Р. [83] |
|
|
Марийцы |
100 117 103 |
0,0 0,0 0,0 |
0,0 1,7 0,0 |
100 98,3 100 |
|
|
Абхазы |
116 |
0,0 |
7,8 |
92,2 |
Саламатина Н.В. и соавт. [62] |
|
Ненцы лесные |
58 129 277 |
3,5 0,0 0,4 |
12,1 Ш 11,5 |
84,5 87,6 88,1 |
Сукерник Р.И. (цит. по Ю. Г. Рычкову и соавт. [13]) |
|
Нганасаны |
235 195 |
0,0 0,0 |
1,3 9,7 |
98,7 90.3 |
|
|
Крс |
Японцы |
5 974 |
0,18 |
[225] |
|
|
Jsa |
Афроамериканцы |
1 298 |
15,8 |
[176,212,3571 |
|
|
Черные африканцы |
593 |
15,6 |
[352] |
||
|
UP |
Финны |
2 620 |
2,6 |
[141] |
|
|
Англичане |
5 000 |
||||
|
Шведы |
501 |
0,2 |
|||
|
Китайцы |
12 |
1 чел. |
|||
|
Японцы |
8 000 |
0,46 |
[300] |
||
|
К17 |
Англичане |
11 044 |
0,29 |
[356] |
|
Столь существенные различия, по-видимому, обусловлены несколькими причинами.
Первая причина - малые или не сопоставимые по величине выборки (см. табл. 5.6, строки 9, 12, 19, 22, 31, 33). Например, у бурят частота антигена К составляет 0,22 %, поэтому обнаружить его в выборке, состоящей из 99 человек, маловероятно (см. табл. 5.6, строка 9).
Вторая причина - артефакты, обусловленные скрытой двойной специфичностью тестовой сыворотки, которая дает завышенные результаты.
Третья причина - чистота выборки. Какая этническая группа подвергалась обследованию: генетически однородная или метисированная. Например, среди алеутов, чукчей, эвенов и эскимосов антиген К встречается чрезвычайно редко. Очевидно, что он представляет собой несвойственный монголоидам признак, привнесенный европеоидами. У жителей некоторых населенных пунктов антиген К не выявляется, у жителей других населенных пунктов - выявляется в единичных случаях лишь у отдельных членов родовых общин. Однако при малой выборке (10-12 человек) даже один позитивный результат дает высокий процент встречаемости. По-видимому, частота антигена К в изоляте составляет 0 %, а в другой, менее изолированной, метисированной выборке - более 10 % (см. табл. 5.6, строки 4,28, 31, 32).
Среди немитисированных с европеоидами северных алтайцев, башкир и якутов фактор К практически отсутствует (см. табл. 5.6, строки 5, 7, 33), тогда как среди метисированных он встречается с частотой 7,3,7,8 и 3,7 % соответственно.
Не исключено, что результаты исследования частоты фактора К могут быть искажены вследствие перекрестного реагирования антигенов и антител системы Kell у представителей разных рас. В частности, тестовые сыворотки, полученные от европеоидов, вероятно, могут в отдельных случаях неадекватно реагировать с эритроцитами представителей монголоидных рас, и наоборот, сыворотки, полученные от монголоидов и негроидов, могут давать с эритроцитами европеоидов позитивные реакции, не имеющие отношения к системе Kell. В результате подобных реакций фактор К ошибочно регистрируют там, где он фактически отсутствует.
Во многих популяциях частота антигенов Kell не исследована. Интересно сравнить эти показатели у народов Дагестана, Сибири, Дальнего Востока, Киргизии, Казахстана и других народов, населяющих территории СНГ.
- Категория: Система Kell
Антиген Km фенотипически тесно связан с антигенами Kell и Кх и так же, как антигены системы Kell, денатурируется при обработке эритроцитов АЕТ. На основании этого антиген Km был отнесен к системе Kell и получил обозначение ISBT KEL20. Однако принадлежность его к системе Kell или Кх окончательно не доказана и остается принятой лишь условно. Основанием для сомнений служат следующие обстоятельства. Если бы антиген Km относился к системе Kell, то он должен был присутствовать на эритроцитах McLeod, как и другие Kell-антигены, хотя бы в небольшом количестве. Однако антиген Km на эритроцитах McLeod не обнаруживали ни методом прямой агглютинации с сывороткой анти-Km, ни с помощью метода адсорбции - элюции. Более того, лица с фенотипом McLeod продуцируют анти-Кт-антитела, что еще раз свидетельствует об отсутствии антигена Km на их эритроцитах. Из указанного выше очевидно, что гликопротеин Kell не является носителем антигена Km, а KEL и Km - разные гены.
Антиген Km отсутствует на эритроцитах К, то есть не представлен на Кх-протеине и, соответственно, не кодируется геном ХК.
Поскольку антиген Km фенотипически явно ассоциирован с антигенами системы Kell и Кх, данные о том, что он отсутствует на гликопротеине Kell и протеине Кх, не находили объяснения. Issitt и Anstee [204] высказали предположение, что при определенных комбинациях белковых компонентов мембраны, кодируемых независимыми локусами, могут появляться новые антигенные детерминанты, не свойственные этим локусам. Так, продукция Kell-гликопротеина, не содержащего вещества Km, контролируется генами хромосомы 7, а продукция Кх-протеина, также не содержащего вещества Km, контролируется геном хромосомы X. Km-антиген появляется, по-видимому, в результате сочетанного эффекта Kx-KEL в процессе формирования комплекса Кх - Kell.
Не исключено существование самостоятельного гена Km, отсутствующего у лиц с фенотипом К или McLeod, однако какие-либо факты в пользу этого предположения отсутствуют.
- Категория: Система Kell
Система Кх
Система Кх включает 1 антиген - Кх (ISBT 019001 или ХК1). Redman и соавт. [326] выделили протеин Кх посредством иммунопреципитации анти-Кх-антителами и определили его мол. массу, составляющую около 37 кДа. Затем было установлено, что Кх-протеин фосфорилируется и паль-митилируется [120, 121], но не гликозилируется [198, 224]. Он представлен в мембране эритроцитов 10 трансмембранными доменами [198], состоящими из 444 аминокислот (рис. 5.5).
Предполагаемая топология Кх-протеина в мембране эритроцита представлена на рис. 5.1. Белок Кх связан дисульфидной связью через Cis 347 с Cis 72 гликопротеина Kell. Терминальные участки Кх-протеина, N и С, погружены в клетку.
По мнению Russo и соавт. [331], комплекс Кх - Kell формируется в эн-доплазматическом ретикулуме и транспортируется к клеточной поверхности. Отсутствие вещества Кх в эритроцитах приводит к уменьшению экспрессии Kell-антигенов на поверхности клеточной мембраны (фенотипу McLeod).
Система Кх кодируется Х-сцепленным геном, ХК, и является самостоятельной антигенной системой независимой от Kell, хотя и тесно с ней связанной.
Кх-протеин выражен на эритроцитах лиц с фенотипом Ко сильнее, чем на эритроцитах с обычным Kell-фенотипом [267]. Однако указанная особенность, как полагают Lee и соавт. [238], Carbonnet и соавт. [122], обусловлена не столько количеством Кх-протеина на этих клетках, сколько тем, что в эритроцитах обычного Kell-фенотипа Кх-протеин связан с белком Kell, что мешает ему с той же силой участвовать в реакции агглютинации с сывороткой анти-Кх.
В противоположность антигенам Kell антиген Кх не денатурируется дити-отрейтолом (ДТТ) и 2-аминоэтилизотиоурониумбромидом (АЕТ), которые разрушают дисульфидные связи. Эритроциты, обработанные ДТТ и АЕТ, приобретают серологические свойства эритроцитов Ко с высокой степенью экспрессии Кх-антигена [102,111].
Анти-Кх
У пациента Klaas, имевшего фенотип McLeod (см. Антиген KL), развилась гемолитическая трансфузионная реакция, вызванная антителами анти-KL, которые, как вскоре выяснилось, представляют собой смесь двух антител разной специфичности и могут быть отделены друг от друга с помощью адсорбции и последующей элюции (Van der Hart и соавт. [377], Marsh и соавт. [258, 267]). Антигены, выявляемые с помощью этих антител, получили обозначение Кх и Km, а соответствующие им антитела - анти-Кх и Km.
Анти-Кх-антитела, как установили Marsh и соавт. [267], сильно реагируют с эритроцитами Ко, но не реагируют с эритроцитами McLeod
С эритроцитами обычного Kell-фенотипа они взаимодействуют слабо. Таким образом, антиген Кх присутствует на эритроцитах Ко, отсутствует на эритроцитах McLeod, выражен, хотя и слабо, на эритроцитах лиц, имеющих нормальный Kell-фенотип.
Таблица 5.5
Выраженность антигенов Кх, Km и Kell на эритроцитах лиц с нормальным и Kell-дефицитными фенотипами
|
Фенотип |
Выраженность антигена |
||
|
Кх |
Km |
антигены Kell |
|
|
Обычный |
+ |
++++ |
++++ |
|
McLeod |
— |
— |
+ |
|
К |
++++ |
— |
— |
|
^"mod |
++++ |
Нет данных |
+ |
« + » - антиген слабо выражен;
« ++++ » - антиген сильно выражен;
« - » - антиген отсутствует.
Анти-Кт-антитела, как установили те же авторы [267], реагируют с эритроцитами обычного Kell-фенотипа, но не реагируют с эритроцитами К и McLeod. Считается (Daniels [141]), что антиген Km формируется в том случае, если на поверхности эритроцита экспрессированы оба белка, Кх и Kell. Если один из них редуцирован, антиген Km отсутствует. Похожая ситуация имеет место в системе Rh-Hr (эффект г/иопозиции). Если гены с и Е наследуются в положении цис, помимо антигенов с и Е вырабатывается сопутствующий антиген сЕ. Этот специфический антиген не является смесью антигенов с и Е. Сыворотки анти-сЕ не реагируют ни с антигеном с, ни с антигеном Е в эритроцитах лиц СЕ/се и се/се, но реагируют с антигеном сЕ в эритроцитах лиц cE/cef где с и Е расположены на одной хромосоме. При наследовании указанных генов в положении транс антиген сЕ не вырабатывается.
Антителаити-1^|+ анти-Кт (анти-KL) вырабатываются у лиц с фенотипом McI||o& Анти-Кт-антитела могут вырабатываться также у гомозигот К°/К°, которые не содержат антигена Km. Причиной появления антител являются трансфузии эритроцитов [114, 177, 267, 377], однако известны антитела нетрансфузи-онного происхождения. Carstairs и соавт. [123] привели случай обнаружения таких антител у пациента, не получавшего трансфузий. Антитела у него выработались после перенесенного септического шока.
Антитела анти-Кх и анти-Km могут вырабатываться как вместе, так и по отдельности. White и соавт. [395] описали пациента с McLeod-синдромом, который выработал только анти-Кт-антитела. Кх-антитела в его сыворотке отсутствовали.
Аналогичный случай привели Sharp и соавт. [345]. У больного имелись анти-Кт-антитела, в связи с чем ему потребовался индивидуальный подбор крови. После пяти трансфузий эритроцитов Ко и Kmod, содержащих сильный Кх-антиген, антитела анти-Кх у больного, вопреки ожиданию, не выработались. Авторы отмечают хороший терапевтический эффект трансфузий. Реакцией отсутствовали.
Russo и соавт. [332] нашли анти-Кх-антитела у пациента с McLeod-синдромом, которому в связи с анемией, развившейся вследствие желудочно-кишечного кровотечения, перелили 4 дозы эритроцитов.
Sullivan и соавт. [358] обнаружили анти-Кх-антитела у больного, имевшего обычный Kell-фенотип. Антитела оказались клинически не значимыми и, по-видимому, имели аутоиммунную природу. Они не гемолизировали ни собственные эритроциты пациента, ни перелитые ему эритроциты, содержащие антиген Кх.
Возможная роль антител анти-Кх и анти-Km в развитии ГБН остается для исследователей невыясненной, поскольку эти антитела у женщин не встречаются, а содержатся только у мужчин.

- Категория: Система Kell
К13-эффект
К13-эффект правильнее называть эффектом отсутствия гена К13, поскольку гаплотип К13-, а не гаплотип К13, содержит ген, подавляющий экспрессию других антигенов системы Kell и napa-Kell (см. К13).
Marsh и соавт. [262], впервые обнаружившие человека с фенотипом К13-, вначале полагали, что низкая экспрессия антигенов k, Kpb, Jsb, Ku и К12 на эритроцитах этого человека связана с его гетерозиготностью по гену К°. Эритроциты пробанда сильно реагировали с сывороткой анти-Кх, что свидетельствовало о наличии у этого человека гаплотипа К°. Генотип пробанда соответствовал формуле К13-/К13-К. Однако, как показали Kaita и соавт. [220], гетерозиготность по гену К° не могла быть причиной депрессии других Kell-антигенов, поскольку люди, являющиеся генетически K°/kKpbJsb, нормально экспрессируют антигены k, Крь и Jsb. Таким образом, Marsh и соавт. [262] оставалось сделать единственно правильный вывод относительно ослабления антигенов Kell у лиц К13-, а именно: низкая экспрессия Kell у лиц К13- обусловлена антитетичным партнером К13, который, будучи в гомозиготной форме и в позиции цис к другим генам KEL, кодирует более слабую, чем в норме, экспрессию антигенов Kell.
У людей К13- слабо выражены антигены К18 (Barrasso [98]), К19 и TOU (Sabo и соавт. [337]).
Вполне вероятно, что случаи ослабления Kell-антигенов, наблюдаемые многими авторами, связаны с гетерозиготностью по К13-, однако это пока невозможно доказать, поскольку отсутствуют сыворотки против указанного гипотетического аллеля К13.
Можно также предположить, что К13-эффект никак не связан с антигеном К13, а является проявлением пока неизвестного генетического процесса, приводящего к уменьшению синтеза антигенов Kell, в том числе антигена К13, который в этом случае не производится вовсе и его экспрессия соответствует 0.
Ожидаемые, но не встречающиеся гаплотипы KEL
Как известно, групповые антигены крови наследуются кодоминантно и независимо друг от друга. Однако существует много исключений, которые ставят под сомнение непреложность этого правила и еще раз подчеркивают огромное разнообразие не только форм групповых антигенов крови, но и способов их наследственной передачи. Приведем некоторые примеры:
- частичное доминирование (эпистазия) D над С в позиции цис. В серологических реакциях это проявляется следующим образом: эритроциты лиц CDe/cde (С и D цис) медленнее агглютинируются сывороткой анти-С и агглютинация выражена слабее, чем при тестировании эритроцитов лиц Cde/cDe (CnD транс);
- ослабление выраженности антигена D (Du) под действием гена С, расположенного по отношению к гену D в позиции транс. Этот феномен проявляется в том, что эритроциты лиц CDe/cde реагируют с сывороткой анти-D значительно сильнее, чем эритроциты лиц Cde/cDe;
- Кра-эффект - подавление синтеза антигенов Kell геном Кра; -К13-эффект - подавление синтеза антигенов Kell неизвестным геном, присутствующим в гаплотипе К13-.
Следует упомянуть еще об одном феномене - неравновесном сцеплении, которое проявляется в том, что антигены двух групповых систем предпочтительно наследуются вместе. Например, у людей, имеющих группу крови М, антиген S встречается в 2 раза чаще, чем у людей, имеющих группу крови N. Среди лиц М+ 72 % являются S+, среди N+ - 31 % S+. Считается, что ген S близко расположен к гену М, поэтому чаще наследуются вместе с ним, чем с геном N. Неравновесное сцепление генов Ми S по сравнению cNnSпослужило основанием полагать, что антигены MN и Ss представляют собой тесно связанную антигенную систему MNS.
В наследственной передаче антигенов Kell также проявляются свои особенности. Помимо Кра- и К13-эффекта существует еще одна необычная генетическая закономерность, а именно отсутствие Kell-гаплотипов, которые по логике вещей должны присутствовать. Речь идет о гаплотипах, кодирующих два редких антигена Kell и более, например: КраК, KJsa, KpaJsa, ККр° и др. Такие гаплотипы до сих пор не обнаружены.
Фенотипы, несущие два редких антигена, встречаются, например:
K+k+Kp(a+b+), K+k+Js(a+b+), K+k+Ul(a+)Kp(a-b+). Однако во всех случаях семейных исследований [154, 249, 251, 252, 288, 357, 402] установлено, что лица с фенотипом K+k+Kp(a+b+) генетически являются KKpb/kKpa, лица с фенотипом K+k+Js(a+b+) имеют генотип KJsb/kJsa, а лица с фенотипом K+k+Ul(a+)Kp(a-b+)- генотип KKpb/kUla.
Возникает вопрос: почему гены К и Кра, К и Jsa9 К и UIa, а также гены других редких Kell-антигенов не наследуются по два и более в одном гаплотипе?
До настоящего времени стройной генетической концепции, объясняющей это явление, не предложено. Некоторые авторы разделяют точку зрения Chown о том, что kKpbJsb представляет собой эволюционно более ранний гаплотип, который в результате единичных мутаций трансформировался в гаплотипы KKpbJsb, kKpaJsb, kKpcJsb и kKpbJsa. Далее логика нарушается, поскольку остальные 5 редких генов: Ula, Wka, К23, К24 и К25 - не имеют аллелей. Согласно предположению Chown, гаплотипы ККра, KJsa и KpaJsa могли сформироваться вследствие двойных мутаций, вероятность которых ничтожно мала в связи с редкостью генов К, Кра, Крс, Jsanjsp. Однако, как полагают Issitt и Anstee [204], с момента открытия редких антигенов Kell выполнено огромное количество исследований, и если бы такие двойные мутации (мутации предыдущих мутаций) существовали, они были бы обнаружены.
Не лишена оснований и другая точка зрения, что антигены системы Kell кодируются 10 гаплотипами (рис. 5.4). Один из них - общий (частый), 9 других -редкие. Первый гаплотип (общий) состоит из гена R- (R-минус), кодирующего все антигены Kell, за исключением редких. Другие 9 гаплотипов представлены одним из редких генов, которые кодируют 1 редкий и все остальные часто встречающиеся антигены.
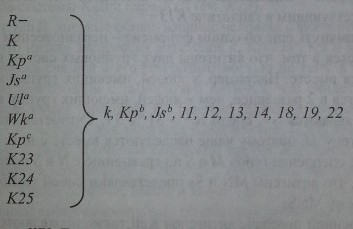
Гаплотипы KEL. Гаплотип 7?- производит общие Kell-антигены: k, Kpb, Jsb, 11, 12, 13,14,18,19 и 22; гаплотип К- антиген К и общие Kell-антигены; гаплотип Кра - антиген Краи общие Kell-антигены и т. д.
В соответствии с этой схемой гаплотипы, содержащие 2 редких антигена и более, исключены. Остается непонятным, почему в локусе KEL отсутствуют генные конверсии и другие формы обмена генетическим материалом между гомологичными хромосомами, как это имеет место в системе резус. По-видимому, в этом проявляется своеобразие рассматриваемого генного локуса.
Итак, подавляющее большинство людей гомозиготны по гаплотипу ШВ и соответственно содержат весь набор общих Kell-антигенов: k Kpb Jsb 11 12 13 14
- Категория: Система Kell
|
Экзон |
Кодон |
Размер, к |
Кодируемые домены и антигены |
|
|
1 |
5'нто Metl |
0,34 |
|
|
|
2 |
2-27 |
0,29 |
Интрацеллюлярный домен |
|
|
а 5 |
28-74 |
0,26 |
Трансмембранный домен |
|
|
4 |
75-133 |
-2,6 |
К18 |
Экстрацеллюлярный домен |
|
5 |
134-175 |
0,33 |
|
|
|
6 |
176-224 |
-3,2 |
К14/К24/К/к |
|
|
7 |
225-245 |
0,093 |
|
|
|
8 |
246-308 |
0,23 |
Кра/Крь/Крс;К11/К17 |
|
|
9 |
309-358 |
-1,3 |
К22 |
|
|
10 |
359^101 |
-6 |
К23 |
|
|
11 |
402-438 |
-1,6 |
тои |
Экстрацеллюлярный домен |
|
12 |
439^171 |
0,24 |
|
|
|
13 |
472-497 |
0,44 |
К19,1Ла |
|
|
14 |
498-531 |
0,19 |
|
|
|
15 |
532-568 |
0,15 |
К12 |
|
|
16 |
569-590 |
0,23 |
HELLH |
|
|
17 |
591-647 |
0,35 |
Jsa/Jsb |
|
|
18 |
648-679 |
-1,3 |
|
|
|
19 |
680-732 3'нто |
|
|
|
Кра-эффект
lien и Lewis [87], открывшие антиген Кра и изучившие характер его наследования, обратили внимание на тот факт, что антиген к слабо выражен у некоторых членов семьи, имевших фенотип Кр(а+). На эритроцитах K-k+wKp(a+b~) антиген к был подавлен до такой степени, что его присутствие могло остаться незамеченным, если бы не были применены сильные анти-к-сыворотки. На эритроцитах K-k+Kp(a-b+) экспрессия к была в нормае.
Объяснение этому феномену нашли Race и Sanger [318], показав, что угнетающее действие гена Кра распространяется не только на к, но и на другие антигены системы Kell.
![Результаты исследования семьи, показывающие депрессивный эффект Кра на k (по Walsh и соавт. [390]). Ослабление к наблюдали у I м (Кра/К°) и П 2 (Кра/К).](/images/stories/stat/ad3.jpg)
Результаты исследования семьи, показывающие депрессивный эффект Кра на k (по Walsh и соавт. [390]). Ослабление к наблюдали у I м (Кра/К°) и П 2 (Кра/К).
В более поздних исследованиях было подтверждено супрессирующее влияние гена Кра на другие гены KEL, расположенные в позиции цис. Опубликованные данные свидетельствуют об уменьшенной экспрессии антигенов Ku (Ford [164]), Jsb (Ford [164], Tippett [370]), K18 (Barrasso и соавт. [98]), K19 (Sabo и соавт. [337]), TOU (Marsh и соавт. [272]) и других часто встречающихся антигенов Kell и napa-Kell (Walsh и соавт. [390]).
Manny и соавт. [256] сообщили, что экспрессия антигена К22 на эритроцитах Кр(а+Ь-) снижена по сравнению с эритроцитами Kp(a-b+). Ваг Shany и соавт. [96], напротив, констатировали, что экспрессия антигена К22 на эритроцитах Кр(а+Ь-) и Кр(а-Ь+) одинакова. Подавляющее действие гена Кра на другие гены KEL не всегда выявляют при обычном серологическом исследовании.
Впоследствии выяснилось, что супрессирующий Кра-эффект можно отчетливо наблюдать только при одном из трех обязательных условий:
- гомозиготность по гену Кра;
- присутствие гена К в позиции транс;
- присутствие молчащего гаплотипа К.
Кра-эффект сильнее проявляется при 3-м варианте, когда лица Кр(а+) имеют генотип Кра/К° [164, 390]. Присутствие гаплотипа К на противоположной хромосоме позволяет более четко увидеть подавляющее действие гена Кра на другие гены KEL в Х/?*-гаплотипе. Подобно этому подавляющее действие гена Кра на экспрессию антигена к и других Kell-антигенов легче обнаружить у людей kKpa/KKpby чем kKpa/kKp\ т. е. когда гены КраиК находятся в положении транс.
На рис. 5.3 представлены результаты семейного исследования, демонстрирующие супрессивный эффект гена Кра на ген к. Ослабление антигена k (к+w) наблюдалось у представителя первого (I м) и второго (II 2) поколения: I м имела генотип Кра/К°, II 2 - генотип Кра/К (Кра транс по отношению к К). У I м все высокочастотные антигены были супрессированы. У других членов семьи антигены Kell имели нормальную выраженность.
Tippett [370] установила, что сыворотки анти-k и aHra-Jsb имели разный титр при титровании 5 образцами эритроцитов Кр(а+Ь-). Один образец эритроцитов от человека k+ Kp(a+b~) Js(a-b+), второй гаплотип которого был, по-видимому, К, показывал отчетливо низкий титр; другие 4 образца k+ Kp(a+b-) Js(a4)+), полученные от гомозигот kKpaJsb/kKpaJsb, показали небольшое снижение титра антител анти-k и aHTH-Jsb, что свидетельствовало о сниженной экспрессии антигенов к и Jsb на эритроцитах гетерозигот К°.
Daniels и соавт. [150], Yazdanbakhsh и соавт. [404] полагают, что Кра-эффект проявляется только низкой синтезирующей способностью Хра-аллеля, в результате чего количество гликопротеинов Kell на эритроцитах снижено. Конформационные изменения в Kell-гликопротеине, которые могли бы препятствовать связыванию Kell-антигенов с антителами, не столь выражены.
Waheed, Kennedy [385] на модели экспрессии кДНК-конструктов в клетках эмбриональной почки человека нашли, что Кра (Тгр 281) замедляет продвижение Kell-гликопротеинов к поверхности клетки, удерживая их на уровне аппарата Гольджи. Для того чтобы обеспечить более эффективное продвижение Kell-гликопротеинов к клеточной поверхности, возникли мутации: часто встречающаяся - Kpb (Arg 281) и редко встречающаяся - Крс (Glu 281).
Daniels и соавт. [150] описали фенотип Ко, который на самом деле таковым не являлся. Несмотря на то что эритроциты этого человека не содержали Kell-антигенов, тем не менее они адсорбировали анти-к-антитела, которые затем обнаруживали в элюате. Кроме того, на эритроцитах пробанда отсутствовал антиген Кх, наличие которого является характерным признаком Ко-фенотипа. Наконец, 3 детей пробанда были Кр(а+Ь+), и маловероятно, что пробанд имел ген К°. Анализ экзона 8 показал, что пробанд был гомозиготен по гену Кр". Авторы отметили также мутации в Ж-гене, что позволило в итоге диагностировать у пробанда фенотип McLeod. Как считают авторы, Kell-ингибирующий эффект, приведший к Ко-подобному фенотипу, был обусловлен двумя причинами: гомозиготностью по гену Краи дефицитом Кх-протеина.
Необходимо отметить, что ^"-эффект проявляется только в отношении часто встречающихся Kell-антигенов. Редкие антигены Kell (имеющие частоту менее 10 %): К, Jsa, Кра, Крс и др. - не подвержены ингибиции, поскольку ген Краи гены К, Jsa, Кра, Крс в одном гаплотипе, в положении цис, не встречаются (см. Ожидаемые, но не встречающиеся гаплотипы KEL).
Kikuchi и соавт. [225] привели данные, показывающие, что в отличие от гена Кра его редко встречающийся аллель Крс не влияет на гены KEL, с которыми он находится в положении цис, как это имеет место при Кра-эффекте. Авторы не отметили подавления экспрессии антигенов k, Jsb и других часто встречающихся антигенов Kell и napa-Kell на эритроцитах лиц с фенотипом Кр(а-Ь-с+).
- Категория: Система Kell
К выводу о том, что локус KEL расположен на хромосоме 7, пришли Zelinski и соавт. [406], Parsons и соавт. [303]. Авторы обнаружили связь между генами KEL и геном пролактининдуцирующего протеина (PIP), который локализован на хромосоме 7 в позиции q32-q36.
Purohit и соавт. [315] нашли, что локус KEL связан с геном цистофиброза, который, так же как PIP, локализован на длинном плече хромосомы 7.
Последующие исследования, проведенные Murphy и соавт. [295], Lee и соавт. [244] с помощью методов гибридизации in situ и метода клонирования Kell-специфической кДНК, подтвердили расположение генов KEL на участке 7q33-7q35.
Генный комплекс Kell включает 19 экзонов и занимает примерно 21,5 кб (Lee и соавт. [242]).
Экзон 1 содержит 5!-нетранслируемую область, кодон метионина 1, инициирующий трансляцию, SP1- и GATA-1 -связывающие участки. Как полагают Camara-Clayette и соавт. [119], экзон 1 вовлечен в отрицательную регуляцию промотора в неэритроидных тканях.
Экзон 2 кодирует цитоплазматический домен и, возможно, второй инициирующий трансляцию метионин 20.
Экзон 3 кодирует трансмембранную, наиболее короткую часть Kell-пожпептида, которая состоит примерно из 20 аминокислотных остатков (см. рис. 5.2).
Экзоны 4-19 ответственны за экстрацеллюлярный, самый протяженный домен. Аминокислотная последовательность, придающая Kell-протеину свойства цинкзависимой металлопротеиназы, кодируется экзоном 16, который, по данным Lee и соавт. [242], на 54,5 % идентичен эквивалентному участку гена NEP, кодирующего вазоконстриктор неприлизин.
Lee и соавт. [242] нашли, что 5'-латеральная область от -176 до -1 нуклеоти-да содержит три участка, связывающих GATA-1. По мнению Shivdasani и Orkin
м. ZU4], GATA-1 является транскрипционным фактором, необходимым для формирования эритроидных тканей.
Мутации в участке связывания GATA-1 приводят к нарушению синтеза на эритроцитах гликопротеина Duffy; эти мутации наблюдают у большинства людей с фенотипом Fy(a-b-). Неизвестно, определяют ли эти мутации отсутствие белка Kell на эритроцитах людей с фенотипом Ко.
Lee и соавт. [242] полагают, что локус KEL регулируется эритроидными факторами транскрипции.
Организация гена KEL*
|
Экзон |
Кодон |
Размер, кб |
Кодируемые домены и антигены |
|
|
1 |
5'нто Metl |
0,34 |
|
|
|
2 |
2-27 |
0,29 |
Интрацеллюлярный домен |
|
|
а 5 |
28-74 |
0,26 |
Трансмембранный домен |
|
|
4 |
75-133 |
-2,6 |
К18 |
Экстрацеллюлярный домен |
|
5 |
134-175 |
0,33 |
|
|
|
6 |
176-224 |
-3,2 |
К14/К24/К/к |
|
|
7 |
225-245 |
0,093 |
|
|
|
8 |
246-308 |
0,23 |
Кра/Крь/Крс;К11/К17 |
|
|
9 |
309-358 |
-1,3 |
К22 |
|
|
10 |
359^101 |
-6 |
К23 |
|
|
11 |
402-438 |
-1,6 |
тои |
Экстрацеллюлярный домен |
|
12 |
439^171 |
0,24 |
|
|
|
13 |
472-497 |
0,44 |
К19,1Ла |
|
|
14 |
498-531 |
0,19 |
|
|
|
15 |
532-568 |
0,15 |
К12 |
|
|
16 |
569-590 |
0,23 |
HELLH |
|
|
17 |
591-647 |
0,35 |
Jsa/Jsb |
|
|
18 |
648-679 |
-1,3 |
|
|
|
19 |
680-732 3'нто |
|
|
|
- Категория: Система Kell
Гены KEL передаются по наследству кодоминантно по тем же законам наследования, как другие групповые признаки крови человека - ABO, MNSs, Rh и т. д. Они не сцеплены с полом (хотя частично зависят от хромосомы X) и не меняются в течение жизни. При отсутствии какого-либо антигена у родителей у детей он также отсутствует.
Присутствие антигенов Kell на эритроцитах (Kell-фенотип) зависит от взаимодействия двух генов. Один из них (аутосомный ген KEL) кодирует синтез полипептида Kell, несущего антигены Kell. Он расположен на хромосоме 7 в позиции q33~35 рядом с локусом пузырного фиброза [244, 295, 315, 406]. Другой ген (ХК) кодирует синтез протеина Кх, который в свою очередь регулирует степень выраженности антигенов Kell на эритроцитах. Этот ген расположен в другом месте - на коротком плече Х-хромосомы, в локусе р21 [103, 200].
Имеются сведения, что ген ХК находится между локусом мышечной дистрофии Дюшена и локусом хронического гранулематоза [103, 200], что позволяет понять причинно-следственную связь указанных заболеваний (гранулематоза и фенотипа McLeod) и предрасположенности к ним мужчин.
Локус KEL, как указывалось выше, включает 5 групп аллельных генов, а также несколько генов napa-KEL, не имеющих аллелей. Пары образуют гены: К/к, Jsa/Jsb, Wka/Wkb и К14/К24. Группа Кра/Крь/Крс содержит 3 аллеля.
Пара-Ке11-антигены экспрессированы на эритроцитах независимо от других антигенов Kell: К, k, Кр, Js и т. д. На эритроцитах McLeod они также слабо выражены, как и Kell-антигены. При фенотипе Ко антигены napa-Kell, так же как и Kell, отсутствуют, что указывает на принадлежность обеих групп к одной системе. Как упоминалось выше, антигены napa-Kell и Kell расположены на одном и том же Kell-протеине мембраны эритроцитов.
Уместно еще раз отметить, что гены К и Кра не могут наследоваться от одного родителя, поскольку никогда не располагаются вместе на одной хромосоме. При исследовании семей не найдено одновременной передачи этих антигенов только от одного из родителей, т. е. гаплотипа ККра [318]. Ген к менее активен, когда передается с геном Кра [318], последний подавляет синтез антигена к (Кра-эффект).
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}