Поиск по сайту
- Вы здесь:
-
Главная

-
Новости
-
Наш блог
-
book
-
Группы крови человека
- Система RH
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
Система RH
Встречаются люди, эритроциты которых не содержат одного, нескольких или всех антигенов Rh или содержат их в редуцированном количестве. До 1980-х годов, когда методы молекулярно-генетического анализа еще не были доступны как сегодня, отсутствие антигенов объясняли выпадением (делецией) соответствующего генетического материала и относили к группе аномалий, получивших название «фенотипы делеции».
Термин не совсем точный, поскольку молекулярные исследования последующих лет показали, что генетический материал у таких людей нередко присутствует, однако не продуцирует Rh-полипептиды.
Отсутствие или слабую выраженность антигенов Rh объясняли также особенностями функционирования гибридных генов, в которых часть экзонов гена D заменена экзонами гена СЕ, и наоборот.
Тем не менее понятие «фенотипы (гаплотипы) делеции» удобно, поскольку объединяет в себе все формы выпадения или редукции Rh-антигенов, которые, несмотря на разное происхождение, имеют одинаковое фенотипиче-ское проявление, то есть не выявляются или слабо выявляются серологическими методами.
Известны следующие фенотипы делеции: -D- [551], D** [236, 243], DCW- [327], Dc- или Dc(e) [649], DIV(C)- [593], Rhnull [346, 347, 689]. Скобками выделяют слабовыявляемые антигены: (е), (С) и т. д. Знак минус означает отсутствие в эритроцитах только антигенов С, с, Е и е, но не других Rh-антигенов как часто встречающихся - Rh total (Rh29), HrB (Bastiaan, Rh34), Rh39, выявляемых аутоантителами, Nou (Rh44), Sec (Rh46), Dav (Rh47), MAR (Rh51), так и редко встречающихся - Cw, Goa, Evans, Таг и др. (табл. 4.22), которые могут присутствовать в этих эритроцитах.
Описано более 70 семей, члены которых были гомо- и гетерозиготными по упомянутым делениям [374, 544]. Чаще такие фенотипы встречались в семьях, где родители были близкими родственниками. Race, Sanger [544] собрали сведения о 23 лицах -D-/-D-, у 20 из них родители состояли в родстве.
Выпадение антигенов обнаруживали при выяснении причин ПТО и ГБН. Лица с фенотипом делеции легко вырабатывают резус-антитела к недостающим у них Rh-антигенам, что и служило причиной осложнений.
Антитела у лиц с Rh-делениями отличаются от резус-антител, обнаруживаемых у резус-отрицательных людей, своей необычностью. Именно в сыворотках этих людей найдены антитела анти-Нго, анти-Hr, анти-Нгв, анти-Nou, анти-Dav и другие, позволившие открыть одноименные антигены. Обычные люди (без Rh-делеций), как правило, содержат все перечисленные часто встречающиеся антигены и не могут вырабатывать к ним антитела.
Многие образцы эритроцитов, относящиеся к фенотипам делеции, за исключением Rhnull, содержат антигены D, G, Rh total и Rh39.
Первый из выявленных фенотипов делеции (-D-) обнаружили Race, Sanger и Selwyn в 1951 г. Затем были описаны другие случаи гомо- и гетерозигот -D-[149, 229, 480, 502, 556, 592, 725]. Последний случай опубликован в 1991 г. Moores и соавт. (Humen Hered., 1991, V. 41, R 295).
Эритроциты -D- не содержат антигенов С, с, Е и е. При этом антигены D, G, Rh total и Rh39 сильно выражены; часто встречающиеся антигены: Hro, Nou, Dav и др. - отсутствуют.
В России фенотип -D- обнаружен в 1985 г. В.А. Мороковым [82] и подробно исследован Т.М. Пискуновой с соавт. [87]. Описанный случай представляет собой классический пример фенотипа -D-.
Наблюдение. Беременная 3., 35 лет, В(Ш) Rh+, диагноз - неразвивающаяся беременность 24 недели. В анамнезе 1 гемотрансфузия и 6 беременностей, из которых первые 2 закончились рождением здоровых детей, 3-я и 4-я - искусственным прерыванием, 5-я - внутриутробной гибелью плода, 6-я беременность, неразвивающаяся, послужила показанием к экстирпации матки. Эритроциты 3. содержали антиген D, но не содержали антигенов С, с, Е, е и Cw.
Бабушка, отец и мать 3. имели гаплотип -D- в гетерозиготном варианте. Всего в семье обнаружено 3 гомозиготы и 6 гетерозигот
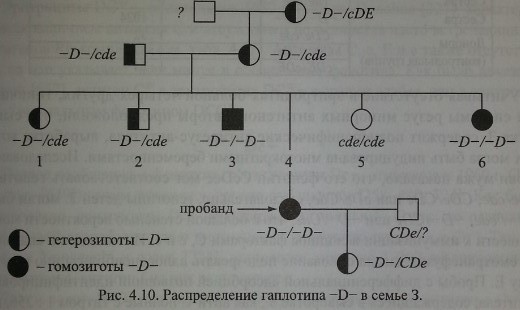
Эритроциты гетеро- и гомозигот -D- проявляли необычайно высокие агшютина-бельные свойства. Титр сывороток анти-D при титровании этими эритроцитами соответствовал 1:256-1:1024, в то время как с эритроцитами доноров контрольной группы титр антител не превышал 1:64 (табл. 4.23). Наиболее выраженная агглютинабельность эритроцитов отмечена у гомозигот -D-/-D-. Результаты этих экспериментов совпадают с данными других авторов, свидетельствующими о том, что эритроциты -D- содержат повышенное количество D-несущих полипептидов (см. Экспрессия антигена D).
Сыворотка 3. реагировала со всеми образцами стандартных эритроцитов независимо от сочетания Rh-антигенов, поэтому подобрать ей совместимую кровь среди доноров оказалось невозможным. С собственными эритроцитами и эритроцитами одного из братьев и одной из сестер сыворотка 3. не реагировала.
В методе солевой агглютинации положительный результат наблюдали только с эритроцитами, содержащими антиген С, что указывало на присутствие в сыворотке полных анти-С-антител.
|
Член семьи |
Генотип |
Титр анти-О-антител с эритроцитами членов семьи и доноров контрольной группы |
|
|
Бабушка |
-D-/cDE |
1 |
: 512 |
|
Мать |
-D-/cde |
1 |
: 512 |
|
Сестра |
-D-/cde |
1 |
:256 |
|
Брат |
-D-/cde |
1 |
256 |
|
Брат |
-D-/-D- |
1 |
1024 |
|
Пробанд |
-D-/-D- |
1 |
1024 |
|
Сестра |
cde/cde |
||
|
Сестра |
-D-/-D- |
1 |
1024 |
|
Доноры (контрольная группа) |
CDe/cde |
1: |
32 |
|
CDe/CDe |
1: |
32 |
|
|
cDE/cDE |
1: |
64 |
|
Учитывая отсутствие в эритроцитах больной четырех других, типичных для системы резус минорных антигенов, авторы предположили, что сыворотка 3. содержит полиспецифические антирезус-антитела, выработка которых могла быть индуцирована многократными беременностями. Исследование крови мужа показало, что его фенотип CcDee мог соответствовать генотипу CDe/cde, CDe/CDe или cDe/Cde, следовательно, генотипы детей 3. могли быть -D-/cde, -D-/CDe или -D-/Cde, что с большой степенью вероятности могло привести к иммунизации женщины факторами С, с и е. Имеющаяся в анамнезе 3. гемотрансфузия давала основание подозревать аллосенсибилизацию к антигену Е. Пробы с дифференциальной адсорбцией позволили идентифицировать антитела, содержащиеся в сыворотке 3., как анти-С полные с титром 1 : 256, неполные анти-с с титром 1 : 2048 и неполные анти-е с титром 1 : 128.
Резюмируя описанное В.А. Мороковым наблюдение, уместно подчеркнуть, что антитела к минорным антигенам С, с и е встречаются относительно редко и имеют низкий титр. На этом фоне способность продуцировать антитела одновременно к обоим антитетичным антигенам - С и с - и к антигену е (hr") в столь высоком титре подчеркивает особый иммунологический статус гомозигот —D -/-/)-, характеризующийся повышенной чувствительностью к недостающим факторам Rh, которые являются для них сильным иммуногеном.
Эритроциты *D* обнаружены Contreras и соавт. [237]. Они содержат повышенное количество антигена Бив серологических реакциях проявляют себя подобно эритроцитам -D-, однако в отличие от последних содержат антигены Evans (Rh37) и Dav (Rh47). Особенности фенотипа *D*, отличающие его от других фенотипов делеций, представлены в табл. 4.22.
Фенотип *D*, как и другие фенотипы Rh-делеций, не содержит общих, широко распространенных Rh-антигенов, присущих нормальному Rh-комплексу (Hro, HrB, Sec и MAR), а также большинства редких Rh-антигенов (Go8, Tar, FPTT и др.).
ИрМиссис Helen Dav, у которой были найдены эритроциты *D*, имела антитела, реагирующие с некоторыми образцами эритроцитов от лиц с Rh-делециями, а ее эритроциты реагировали с некоторыми сыворотками от указанных лиц. Анализ особенностей реагирования эритроцитов миссис Dav привел к открытию одноименного антигена (см. Dav).
DCW-
Эритроциты DCW-, описанные в 1957 г. Gunson, Donohue [327], характеризуются наличием антигена G и отсутствием большинства часто встречающихся антигенов Rh-комплекса (см. табл. 4.22). Антигены С, с, Е и е также отсутствуют, на что указывает знак минус в обозначении фенотипа. Как было показано Tippett [657], антиген Cw в эритроцитах DCW- выражен слабее, чем в эритроцитах с нормальным фенотипом DCCwe.
Dc-
Первое сообщение о фенотипе, обозначенном Dc-, относится к 1960 г. (Tate и соавт. [649]), затем появились другие публикации [258, 440, 724]. Как было установлено, эритроциты Dc- не содержат антигенов С, Е и е, но отличаются большой вариабельностью в отношении количества антигена с (hr'). Так, по данным Yamaguchi и соавт. [724], экспрессия антигена с (hr') на эритроцитах Dc- снижена, антиген f на них отсутствует.
Tate и соавт. [649] нашли, что, несмотря на ослабленный антиген с (hr') и отсутствие антигена е (hr"), эритроциты Dc- содержат антиген f, но в ослабленной форме. Отдельные образцы эритроцитов Dc- содержали некоторое количество антигена е (hr"). Такой фенотип более соответствовал написанию Dc(e), т. е. Dc со слабым е (hr"), или Dc((e)), т. е. Dc с исчезающим количеством е (hr").
Leyshon и соавт. [440] привели данные о нормальной выраженности антигена с (hr') и f (се) на эритроцитах Dc-. Однако Spielman и соавт. [Vox Sang., 1974, V. 27, R 473] описали случай ГБН, вызванный aHTH-f-антителами, у женщины, которая имела генотип CDe/Dc-, что вступает в противоречие с данными Leyshon и соавт.
Многие авторы сходятся во мнении, что количество антигена D в эритроцитах Dc- выше, чем в обычных эритроцитах D+.
Tessel и соавт. [650] описали фенотип Dc- негра и негритянки, не связанных родством. Эритроциты мистера J.W. содержали повышенное количество антигена D, уменьшенную по сравнению с нормой дозу антигенов с (hr1) и f, а также слабый антиген е (hr"). Эритроциты миссис F.J. «держали повышенное количество антигена D, уменьшенную дозу антигена с (hr'), но не содержали антигенов f и е (hr"). Эритроциты мистера J.W. реагировали с антителами анти-Нго/Нг, имевшимися в сыворотке крови миссис F.J. и двух гомозигот -D-/-D-, а также с двумя из трех сывороток анти-nl. Эритроциты миссис F.J. не реагировали с сыворотками анти-Нго/Нг и анти-nl. Авторы пришли к выводу, что ген Dc- гетерогенен и может кодировать 2 фенотипа Dc-, один из которых содержит антиген f, трудновы-являемый антиген е, антигены Нгои Нг, а другой фенотип Dc- не содержит перечисленных антигенов.
DIV(C)-
В 1969 г. группой исследователей [593], включая классиков современной им-муносерологии Sanger, Tippett и Salmon, был описан единственный известный до настоящего времени человек с генотипом DIV(C)-/DIV(C)-. Людей, гетерозиготных по гаплотипу DIV(Q- обнаружено несколько. Гаплотип DIV(C)- производит парциальный антиген D категории IVa, антиген G и значительно уменьшенное количество антигена С.
В гомо- и гетерозиготном варианте генный комплекс DIV(C)- обеспечивает экспрессию редко встречающихся антигенов: Goa(Rh30) [593], RoHar(Rh33), Riv (Rh45) и FPTT (Rh50) [257], а также часто встречающихся Rh-антигенов: Rh total (Rh29), Rh ауто-С-подобный (Rh39), Dav (Rh47) и Nou (Rh44), которые не кодируются другими гаплотипами Rh-делеций (см. табл. 4.22).
Антиген Nou был впервые обнаружен на эритроцитах гомо- и гетерозигот DIV(C)-(cu.Nou).
Хотя эритроциты DIV(C)- лишены антигена е (hr"), 3 из 10 сывороток анти-е давали реакции с этими эритроцитами, что указывает на продукцию некоторого количества антигена е (hr") или Се (rh}) гаплотипом DIV(C) —.
Rhnilll
Среди упомянутых выше фенотипов делеций (-D-, Dc-, DIV(C)~ и DCW-), при которых наблюдается выпадение 2 антигенов и более, фенотип Rhnu|| занимает особое место. Он отличается отсутствием всех антигенов системы Rh-Hr, а также антигенов системы LW и Duffy, кодируемых генами, близко расположенными к локусу RH.
О первой находке необычного генотипау аборигенки Австралии сообщили в 1961 г. Vos и соавт. [689]. В 1964 г. такой же генотип описали Levine и соавт. [428] у белой американки. Двумя годами позже, в 1966 г., Ishimori, Hasekura [371] нашли нулевой Rh-фенотип у японского мальчика.
По данным Race и Sanger [544], к 1975 г. было описано более 20 лиц Rhnull из 14 семей, отличавшихся расовой принадлежностью (табл. 4.24). К этой сводке можно добавить 11 случаев Rhnull, приведенных Daniels [244].
Issitt и Anstee [374] насчитали 33 случая Rhnull, 1 из которых обнаружен ими.
Фенотип Rhn „ у лиц, внесенных в табл. 4.24, диагностирован при различных обстоятельствах. В 1-м случае он был обнаружен при проведении популяцион-ных исследований у австралийских аборигенов. В других случаях (2, 4, 6, 8, 10, 11 и 12-м) - при выяснении специфичности антител, в том числе у больных тяжелой формой гемолитической анемии (8, 11 и 12-м), а также при обследовании доноров, рожениц, новорожденных (3,5,7,9,13 и 14-м).
Родственники носителей Rhnulib некоторых случаях также имели нулевой фенотип или сниженную экспрессию Rh-антигенов. По сводке Daniels [244], из 11 лиц, имевших дефицит Rh-антигенов, 8 были детьми от близкородственных браков. Из общего числа обследованных 6 человек японцы.
Относительно большое число лиц Rhnull среди японцев обусловлено, скорее всего, не тем, что в этой популяции высокая частота указанного фенотипа, а проведением широких популяционно-скрининговых исследований, в процессе которых возможно выявить людей со столь редким фенотипом.
Обозначение--------- /--------- , использованное Vos и Levine с соавторами [428,689], было признано неточным, поскольку означает отсутствие генетического материала у носителей этого фенотипа. В действительности гены RH у таких людей часто присутствуют, но не проявляют себя фенотипически. По предложению Ceppellini фенотип с отсутствием Rh-антигенов получил название Rhnull. Обозначение-------- /-------- сохранилось для описания гена г=, который, как позднее выяснилось, представляет собой молчащий (не производящий продукта) аллель локуса RH.
Отождествление антигенов VS и es на основании только лишь серологического сходства некоторые исследователи считают недостаточно аргументированным.
Оригинальную интерпретацию связи VS и es предложили Steers и соавт. [626], которые установили, что ген RHCE лиц VS+ имеет мутацию, приводящую к замене в Rh-полипептиде лейцина на валин в позиции 245. Поскольку валин в позиции 245 располагается внутри мембраны, можно полагать, что он не принимает непосредственного участия в серологической реакции с анти-VS-антителами, но его присутствие, вероятно, влияет на структуру экзофациальной части полипептида Ее. Валин 245 расположен недалеко от позиции 226, в которой замена пролина на аланин определяет соответственно специфичность Е или е. Steers и соавт. предположили, что специфичность антигена VS обусловлена валином 245, присутствие которого одновременно с аланином 226 способствует трансформации е в es. Таким образом, VS и es, по-видимому, вместе присутствуют на эритроцитах VS+, и далеко не факт, что они являются одним и тем же антигеном.
Faas и соавт. [278] подтвердили данные Steers и соавт. [626], показав, что присутствие антигена VS на эритроцитах связано с заменой лейцина 245 на валин. Авторы предположили, что эта замена может также влиять на экспрессию антигена С, то есть воздействовать на серии в позиции 103, в силу чего образуется фенотип Cdes со слабым антигеном С, ассоциированным с VS+es+. Эти взгляды нашли подтверждение в экспериментах Reid и соавт. [Transfusion, 1995, v. 35 (sappl. 10s), 51s], показавших, что эритроциты Cde, не содержащие антигена hrB (парциального е), часто являются es+. Так, из 48 выявленных образцов эритроцитов hrB- 45 были VS+. Авторы предположили антитетичную взаимосвязь между антигенами VS и hrB. Если VS и hrB кодируются двумя аллелями, то должен существовать третий аллель, который их не образует. Действительно, Reid и соавт. обнаружили образцы эритроцитов VS+hrB-, VS~hrB+, а также редкие образцы VS-hrB-.
Не исключено также, что антиген VS обнаруживают на эритроцитах, когда недостающие эпитопы hrB имитируют антиген es, в то время как в действительности es отсутствует.
Возможно генетическая структура, обусловливающая синтез антигена VS, часто, но не всегда, ингибирует продукцию антигена hrB.
Ces(Rh42)
Щ 1980 г. Moulds и соавт. [493] описали антитела, которые реагировали с эритроцитами Cdes, но не реагировали с эритроцитами Cde, а также с эритроцитами VS (es)+ и V (ces)+, не содержавшими антигена С.
Антитела получили название анти-Се8 по аналогии с анти-Се, поскольку реагировали с антигеном Ces, но не связывались с антигенами С и es, представленными в эритроцитах по отдельности.
Гаплотип Cdes, кодирующий выработку антигена Ces, встречается у негров [546, 574,575,639], в частности у негров банту [730,731]. У белых он редок, Щ 1 %.
Тот факт, что эритроциты негров Cde/cde реагировали не со всеми сыворотками анти-С, сразу же привлек к себе внимание. В 1959-1960 гг. Sturgeon и соавт. [639, 642] установили, что эритроциты большинства негров Cde/cde реагируют с анти-С, но не реагируют, вопреки ожиданиям, с сывороткой анти-Се.
Zoutendyk и Teodorsuk [730, 731] наблюдали такие же необычные Cde/cde-генотипы у представителей южно-африканской народности банту, тестируя их эритроциты сыворотками анти-С и анти-Се.
Поскольку новый фенотип был выявлен среди лиц rf (Cde), соответствующий ему генный комплекс обозначили как r's (Cdes).
0/е5-гаплотип отличается от Cde-гаплотипа тем, что производит антиген esвместо антигена е, хотя частичная продукция антигена е этим гаплотипом сохранена. Напомним, что антиген es не реагирует с анти-е-антителами, а антиген е не дает реакции с анти-е-антителами. Таким образом, антиген es отличается от антигена е. Этим и объяснялось отсутствие агглютинации эритроцитов Cdesсыворотками анти-Се.
Далее выяснилось, что гаплотип Cdes производит антигены f (се) [546, 574, 575, 639] и С° [373], но, как уже упоминалось, не производит антигена rh.(Ce). Об этом свидетельствуют эксперименты, показавшие, что эритроциты Cdes/cde, имеющие г/ис(антигены с и е, а также эритроциты Cdes/cDE, не имеющие цис-антигенов сие, реагируют с сывороткой анти-f (се), дают слабые реакции с сывороткой анти-е [575] и анти-G [373], но вместе с тем не реагируют с сывороткой анти-Се.
Rosenfield и соавт. [574] отметили, что эритроциты лиц Cdes/CDe реагируют с анти-с-сыворотками, что указывает на способность гаплотипа Cdes экспресси-ровать определенное количество антигена hr? (с).
Лица Cdes содержат необычную форму антигена С, реагирующую не со всеми сыворотками анти-С, и не содержат антигена rh. (Се), вследствие чего могут продуцировать антитела анти-С и анти-rh. (Се) [572,639].
Некоторые авторы полагают, что анти-Се 8-антитела в действительности имеют специфичность анти-С Ges, так как эритроциты Cdes, с которыми они реагируют, несут больше антигена CG, чем антигена С. Это указывает на возможное происхождение антигена Ces от гена RHD, а не от RHCE. В действительности фенотип Cdes является продуктом гибридного гена Cdes, одна половина которого состоит из экзонов RHD-rem, а другая - из экзонов RHCE-тет.
Антитела анти-Сеs отличаются от анти-С, анти-е, анти-Се, анти-е8 и других известных Rh-антител, что позволило сделать вывод о существовании еще одного антигена в системе Rh, получившего обозначение Rh42.
Антиген Ces (Rh42) обнаруживают у лиц с гаплотипом Cdes и CDes, в основном у негроидов.
Мнения исследователей относительно обозначения гаплотипа Cdes (как и CDe5) разделились. Одни авторы полагали, что гаплотип, продуцирующий необычную форму антигена С и антиген с, представляет собой Ccdes [544]. Другие предложили обозначать этот гаплотип как cdesCG, подчеркивая тем самым, что антиген С° отличается от антигена С [373].
Определенную ясность в неразрешимую серологическими методами ситуацию внесли молекулярно-генетические исследования Blunt, Daniels, Carritt [174], которые показали, что лица, имеющие гаплотип Cdes и по существу относящиеся к Rh-, содержат экзоны 1, 2, 8, 9 и 10, характерные для гена RHD. Более того, авторы не обнаружили экзона, который мог бы-кодировать антиген С. На этом основании высказано предположение, что атипичный антиген С, или CG, у лиц с гаплотипом Cdes может являться продуктом оставшейся части гена RHD, подвергшегося неполной делеции.
Как уже указывалось, эритроциты Cdes, так же как CDes, не содержат антигена rh. (Се), но имеют антиген f (се). Однако эта особенность присуща только негроидам. У европеоидов, наоборот, эритроциты CDes имеют гЦ (Се)-антиген, но не содержат f (се)-антигена.
Ce-like
Svoboda и соавт. [647] в 1981 г. обнаружили антитела, напоминающие по специфичности анти-Се и анти-Се8. Выявляемый с их помощью антиген получил наименование rh-like и номер Rh41. Антитела affra-Rh41 ведут себя как анти-rh (Се) в реакциях с эритроцитами лиц Cde/cde, однако по по разному реагируют с эритроцитами CwCGcDefrh;CwCcDefrh Описаны единичные случаи антител анти-Ce-like (affra-Rh41). Антитела анти-Сеs (aHra-Rh42) также описаны в единичных случаях. По сравнению с этими двумя специфичностями анти-Се-антитела встречаются значительно чаще, и новые случаи обнаружения этих антител уже не привлекают столь пристального внимания как в 70-80-е годы.
В 1955 г. De Natale и соавт. [259] обнаружили у реципиента cde/cde, имевшего множественные гемотрансфузии, антитела, которые были названы авторами анти-V. Вскоре Giblett, Chase, Motulsky [301] описали еще двоих людей, в сыворотке которых содержались анти-У-антитела, обусловленные, как и в первом случае, переливаниями крови.
Антитела анти-V проявляли гетерогенность при исследовании крови людей, имевших парциальный е-антиген и одновременно содержащих анти-е-подобные антитела.
Антиген V был определенно связан с системой Rh, поскольку всегда встречался у лиц с гаплотипом cde или cDe, то есть имевших гены се в положении цис. Race и Sanger [544] предположили, что ген Vявляется аллелем/(се). После открытия антител анти-VS (анти-е8), реагирующих с образцами эритроцитов V+ и Cdes+, аллельная связь генов Vnf (се) стала очевидной, и антиген V, исходя из результатов серологических реакций, получил наименование ces.
Поскольку антиген V (ces) формируется, когда антиген е (hr") кодируется аллелем es, считалось, что антиген V (ces) соотносится с антигеном es, как антиген f (се) с антигеном е (hr"). В настоящее время многие результаты проверочных серологических исследований совпадают с этим толкованием, однако отдельные данные ставят его под сомнение.
Антиген V встречается преимущественно у негров (табл. 4.20). Среди американских негров и негров Африки, в том числе банту, этот антиген наблюдали с частотой до 40 %. Причем африканские негры, у которых генный комплекс се и ^регистрируют чаще, чем у американских негров, имеют более высокую частоту антигена V. Антиген V чаще присутствует у людей с фенотипом cDe, чем cde. У европеоидов и монголоидов этот антиген бывает редко (0,19-0,49 % и 0,37-1,15 % соответственно).
Shapiro [605], исследуя большое количество образцов крови народа банту в Южной Африке, сделал вывод, что у банту существуют 2 серологически различающихся антигена. Один из них (hrv) по серологическим свойствам идентичен антигену V, описанному DeNatale и соавт. [259]. Другой антиген (hrH -сокращение от Hermanez), получивший номер Rh28, как впоследствии выяснилось, является новым самостоятельным антигеном системы Rh.
Автор нашел, что hrv и hrH широко распространены у банту. Кроме того, он отметил, что каждый фактор или оба фактора могут продуцироваться различными гаплотипами, но чаще всего cde, Cde и cDe.
Экспрессия антигена V обусловлена заменой G —► Т в нуклеотиде 1006 гена RHCE, что приводит к замещению глицина на цистеин в позиции 336 [247].VS (es) [Rh20]
В 1960 г. Sanger и соавт. [595] обнаружили анти-е-подобные антитела, обозначенные VS по имени женщины-носительницы антител - миссис V. S. Анти-VS-антитела реагировали со всеми эритроцитами V+ (cdes) и большей частью эритроцитов Cdes. В табл. 4.21 представлены данные о характере реагирования антител анти-VS и анти-V, позволяющие заключить, что указанные антитела выявляют разные антигены.
Таблица 4.21
Реакция антител анти-V (ces), анти- VS (es)
|
Гаплотип |
Реакция антител |
|||
|
анти-V (ces) |
анти-VS (es) |
анти-С + анти-Се |
||
|
с одними образцами эритроцитов |
с другими образцами эритроцитов |
|||
|
Cdes |
— |
+ |
+ |
-или± |
|
cdes |
+ |
+ |
— |
— |
|
Cde |
— |
— |
|
+ |
|
cde |
— |
— |
— |
— |
* По Race, Sanger [544].
DeNatale и соавт. [259], Sanger и соавт. [595]) установили, что эритроциты VS+ содержат необычную форму е-антигена, которая была названа es, поскольку анти-VS-антитела реагировали с антигеном es в фенотипах cdes и Cdes. Эритроциты VS+ (es+) реагировали с большинством, но не со всеми образцами анти-е-антител, полученных от лиц Е/Е, а также не со всеми анти-е-подобными (парциальными) антителами, полученными от лиц е+. На основании этих данных сделано заключение, что антиген VS (es) кодируется локусом es, который является аллелем Е и е. В то же время антиген V (ces) кодируется геном ces, являющимся аллелем се.
Антигены VS (es) и V (ces), по-видимому, могут формироваться при участии гена RHCe, а именно его разновидности Ces, поскольку некоторые лица (негроиды) с га-плотипом Cdes имеют эритроциты VS+V+, а другие, с гаплотипом Cde, имеют эри-троциты VS-V-. Описаны редкие феногапы VS-V+и C-V-VS+(Shapiro [605]).
Считается, что экспрессия антигена VS обусловлена заменой С —> G в ну-клеотиде 733 гена RHCE, и это приводит к замещению Leu 245 щ Val в экзо-не 5 [278, 356, 626]. У всех обследованных лиц VS+ наблюдали гибридный ген RHD -CE-D, имеющий в экзоне 5 Val 245 [247].
Daniels и соавт. [Transfusion, 1997, Suppl. 9S, 101S] показали, что у негров фенотип VS+V+ ассоциирован с Val 245 на се-полипептиде, а фенотип VS+V- ассоциирован с Val 245 и Cys 336. Его продукция обусловлена экзоном 3 гибридного гена D-CE.
В серологии антигенов V и VS, так же как и других е-подобных, в том числе парциальных антигенов е, много неясного. У негроидов (американских негров, негров банту) и европеоидов они выражены сильнее, чем у монголоидов (южноамериканских индейцев). Известны комбинированные и переходные формы антигенов V и VS, а также антител анти-V, анти-VS, анти-Ьгу, анти-Ьгн и анти-Ьг^, сепарируемых и несепарируемых, что свидетельствует о полиморфизме этих антигенов и антител и затрудняет их идентификацию серологическими методами. Имеется суждение, что антиген VS (es) соотносится с антигенами V (ces) и hrH, как антиген G соотносится с антигенами D и С. То же самое можно отнести к соответствующим антителам.
Некоторые исследователи отметили, что аллоиммунные антитела анти-D и анти-DC, обладающие высокой активностью, в ряде случаев содержат компонент анти-V и реагируют с эритроцитами C-D-V+ [374]. При детальном исследовании этих случаев почти всегда обнаруживалось, что обладатели антител были иммунизированы (в результате беременности или трансфузии) эритроцитами V-.
Отмечена также некоторая связь антигенов VS и V с парциальными D-антигенами и слабым антигеном Du.
hrH(Rh28)
При исследовании сыворотки миссис V. S. адсорбцией - элюцией было показано, что сыворотка этой женщины содержала моноспецифические антитела анти-VS (Sanger и соавт. [595]). Сыворотки анти-VS, исследованные Shapiro [605], содержали комбинированные антитела анти-Ьгу (или V, или ces) + aHTH-hrH, которые можно было разделить дифференциальной адсорбцией. Другие сыворотки анти-VS, помимо сепарируемых антител анти-Ьгу и анти-11гн, имели несепарируемые антитела, которые были названы airra-hrVH, поскольку определяли общую часть аплютиногена, присутствующего на эритроцитах hrv+ и на эритроцитах hr1**.
Антитела анти-VS легче адсорбируются эритроцитами V-VS+, чем эритроцитами V+VS+, так как последние содержат меньше VS-антигена. Антитела анти-hrH отличались от анти-VS. Они полностью адсорбировались эритроцитами hrH+, но не адсорбировались эритроцитами hrH- фенотипов V-VS+ или V+VS+.
Анти-е-подобные аутоантитела
Описаны анти-е-антитела, которые при первом исследовании принимают за аллоиммунные, хотя они имеют аутоиммунное происхождение (Rosenfield и со-авт. [571]). В редких случаях анти-Ьгв и/или Нго-подобные антитела обнаруживали у пациентов в период, когда экспрессия антигенов hrB и Нго на их эритроцитах была подавлена. Антитела рассматривали как аллоиммунные. Через некоторое, иногда довольно продолжительное время нормальная продукция указанных антигенов восстанавливалась и антитела исчезали, что служило основанием предполагать их аутоиммунную природу.
Значение анти-е-подобных аутоантител в трансфузиологии не выяснено. Однако, когда они присутствуют у больного, ему при необходимости переливают эритроциты, которые не агглютинируются указанными антителами.
Моноклональные анти-е-антитела f
Эритроциты людей Е+е- реагируют с некоторыми моноклональными анти-е- и анти-е-подобными антителами, в то время как поликлональные анти-е-антитела дают с этими эритроцитами отрицательные реакции. Пока нет ясности: выявляют ли моноклональные анти-е-антитела какие-то дополнительные эпитопы е, не выявляемые человеческими поликлональными сыворотками, или же различия в реактивности моно- и поликлональных антител обусловлены неодинаковой авидностью?
Как известно, полипептид, несущий антиген е (hrM), имеет в позиции 226 аланин [496]. С такой структурой реагируют как поли-, так и моноклональные анти-е-антитела. Полипептид Rh, несущий антиген Е (rh"), имеет в позиции 226 пролин. С такой структурой антитела анти-е не реагируют. Для того чтобы оценить своеобразие этих взаимодействий, целесообразно упомянуть, что большинство полипептидов D также имеет аланин в позиции 226, однако эта структура, вопреки ожиданиям, не реагирует с поликлональными и большинством моноклональных анти-е-антител и их не адсорбирует.
Высказано предположение, что поликлональные анти-е-антитела и некоторые моноклональные распознают аланин 226, кодируемый геном RHCe и геном RHce, но не геном RHD. В то же время другие моноклональные анти-е-антитела, реагирующие с эритроцитами D+E+e-, могут распознавать аланин 226, кодируемый генами RHCe и RHce, а также геном RHD.
Однако это предположение не нашло подтверждения в экспериментах, показавших, что моноклональные анти-е-антитела не реагируют с полипептидами ~D-, которые имеют аланин 226, но лишены е-антигена
Анти-е-подобные антитела относят к клинически среднезначимым. Введение лицам, содержащим эти антитела, небольших количеств (аликвот) несовместимых эритроцитов приводило к их выведению из кровяного русла. При этом клиренс несовместимых эритроцитов иногда был слабо выражен, что свидетельствовало о вялотекущем их отторжении.
Подбор совместимой крови больным, у которых имеются парциальные анти-е-антитела, не представляет большой сложности, если его проводят из регистра типированных доноров. Эритроциты Е/Е и некоторые е/е (примерно 10 %) являются совместимыми и нормально выживают in vivo.
Если у реципиента имеются комбинированные антитела - парциальные анти-е + парциальные анти-Нго - совместимую кровь подобрать сложно. Подходящими могут оказаться эритроциты с некоторыми делециями Rh.
В случае, если реципиент содержит антитела анти-Нго + анти-Hr, найти совместимую кровь будет крайне сложно. Совместимыми в этом случае могут оказаться лишь уникальные образцы эритроцитов. Теоретически такие ситуации возможны, но они представляют большую редкость и в литературе не описаны. За 45 лет истории ГНЦ (до 2010 г.) наблюдали 1 случай редкой анти-е-подобной сенсибилизации реципиента, которому, однако, удалось совместными усилиями нескольких СПК подобрать совместимую кровь, отобранную в результате исследования образцов крови доноров Е/Е.
По данным других авторов [374], совместимую кровь для реципиентов, имеющих анти-е-подобные антитела, удавалось подобрать с частотой 1-2 донора на 100 обследованных. При наличии анти-Нг -антител вероятность подбора была ниже, но можно было иногда найти совместимые дозы крови. При этом авторы подчеркивают, что большинство людей, у которых образовались эти антитела, были неграми, и найти совместимую кровь среди доноров негров было намного проще.
Имеется несколько сообщений о том, что переливание эритроцитов cDE-фенотипа пациентам, имевшим анти-е- и анти-е-подобные антитела, проходи-9 без немедленной или отсроченной трансфузионной реакции и не приводило к повышению у реципиентов авидности и титра анти-е-антител в посттрансфу-зионном периоде.
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}