Поиск по сайту
- Вы здесь:
-
Главная

-
book
-
Группы крови человека
- Системы Lutheran и Lewis
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...

- Категория: Системы Lutheran и Lewis
Считается, что адсорбция олигосахаридов Lewis на клетках пассивная [115], однако механизм, по-видимому, не столь прост. Транспортная функция эритроцитов изучена мало, хотя очевидно, что перенос многих биологически активных веществ - гормонов, ферментов, вирусов - это сложный микрофизиологический процесс, в котором немаловажная роль принадлежит серологически выявляемым структурам эритроцитов, в том числе Lewis.
Антигены Lea и Lex принимают участие в клеточной дифференцировке и опухолевой трансформации [75, 98] и рассматриваются как фактор, способствующий гематогенному метастазированию опухолевых клеток [97, 153, 222, 241]. Наряду с другими онкоэмбриональными антигенами антиген Lex, по мнению некоторых исследователей, может служить маркером прогрессирующей малигнизации [75, 83, 96, 99].
Lex хорошо выражен в эмбриональных тканях [83, 212] - отсюда его обозначение SSE-1 (Stage Specific Embryotic Antigen-1). Он максимально экспрес-сирован на стадии морулы у мышей и, как полагают [212], играет определенную роль в преимплантации эмбриона.
Fukushi и соавт. [78], исследуя человеческие эмбрионы в разные сроки развития, установили, что Lex появляется через 40 дней с момента оплодотворения, достигает максимума к 50-70 дням, далее его экспрессия уменьшается.
У взрослых Lex присутствует на нейтрофилах, эпителии проксимальных канальцев почек [49, 76, 78] и желудочно-кишечного тракта [200].
Lea, Lex и их производные (Leb и Ley), обработанные сиалидазой, накапливаются в опухолевых клетках, которые в процессе малигнизации утрачивают нормальную дифференцировку [46, 79, 112, 200] и возвращаются к экспрессии антигена Lex, свойственного эмбриональным тканям.
Как показали серии исследований [34, 38, 131, 231], антигены Lex и Lea, обработанные сиалидазой и сульфатазой, являются лигандами адгезивных молекул, обеспечивающих хоминг лимфоцитов при остром воспалении.
Адгезия, так же как и трансэндотелиальная миграция лимфоцитов, обусловлена адгезивными молекулами, экспрессированными на поверхности эндотели-альных клеток [30,147,152].
Антигены Lex, Ley (в меньшей степени Lea, Leb и Led) обнаружены в грамо-трицательныех бактериях Helicobacter pylori [24, 25, 159, 174], вызывающих хронический гастрит [237], язву желудка и двенадцатиперстной кишки [85], лимфому [190] и аденокарциному желудка [181,182, 189].
По мнению Appelmelk и соавт. [20], патогенез Helicobacter-шщуцщюванноп) гастрита включает аутоиммунный компонент и сводится к следующему. Антигенная мимикрия по Lex позволяет Helicobacter pylori, оставаясь незамеченными, нарабатывать большое количество антигена Lex, к которому в итоге вырабатываются нейтрализующие его антитела airra-Lex. Последние, будучи перекрестно реагирующими, взаимодействуют не только с Helicobacter, но и с эпителием желудка, провоцируя гастрит.
Teneberg и соавт. [226] и другие авторы [167,168] полагают, что Helicobacter pylori содержат нейтрофилактивные протеины, способствующие накоплению нейтрофилов в слизистой оболочке желудка.
Щ Другие исследователи [37, 73, 113] связывают роль Helicobacter в этиологии гастрита с непосредственным их прилипанием к слизистой оболочке желудка с помощью адгезивных Ьеь-молекул, что приводит к локальному воспалению.
Молекулярная мимикрия, по мнению ряда авторов [64, 133, 215], лежит в основе заражения человека кровяным гельминтом Schistosoma mansoni, который синтезирует гликоконъюгаты, несущие антиген Lex. Анти-Ьех-антитела, вырабатывающиеся у инвазированных людей, предотвращают повторное заражение [50, 65]. Однако эти антитела способны вызывать комплементзависимый цитолиз гра-нулоцитов, что приводит к нейтропении, наблюдаемой при шистозоматозе [209].
- Категория: Системы Lutheran и Lewis
Слабый (Ье™)-фенотип
Слабая выраженность антигенов Lewis обусловлена аминокислотными заменами в генах, контролирующих продукцию фукозилтрансфераз [71,134,172,180].
Koda и соавт. [134] нашли у японцев Le(a-b-) 2 мутации в гене FUT3 (Leu 20 —> Arg, Gly 170 —> Ser). Опыты с клонированием гена Le в COS-клетках показали, что замена Gly 170 —> Ser приводит к продукции неактивного фермента, тогда как при замене Leu 20 щ Arg вырабатывалась активная фукозилтрансфераза, о чем свидетельствовало появление антигена Leb на поверхности COS-клеток.
Mollicone и соавт. [172] также выявили мутацию Leu 20 щ Arg у индонезийцев Le(a-b-), которые содержали антигены Lewis в слюне. Такие же замены описаны у японцев [180] и шведов [71]. Замена аргинина на лицин приводила к экспрессии слабовыраженного антигена Lewis.
Nishihara и соавт. [179] и Mollicone и соавт. [172] описали мутацию Не 356 —» Lys. Гетерозигота по этой мутации, 18 из 19 индонезийцев Le(a-b-), не содержали антигенов Lewis ни на эритроцитах, ни в слюне.
Cooling и Gu [59] исследовали с помощью ПЦР 15 афроамериканцев, имевших фенотип Lenull, Отмечены варианты мутаций в FUT3, которые приводили к продукции неактивного фермента и в результате формировали нулевой фенотип. Одну из мутаций (G —> А) в нуклеотиде 13 обнаружили у 50 % обследованных (рис. 9.3), она приводила к замене Gly 5 —£ Ser. У других обследованных обнаружили мутации G —► Т в нуклеотиде 1022, G Щ А в нуклеотиде 484, G ->
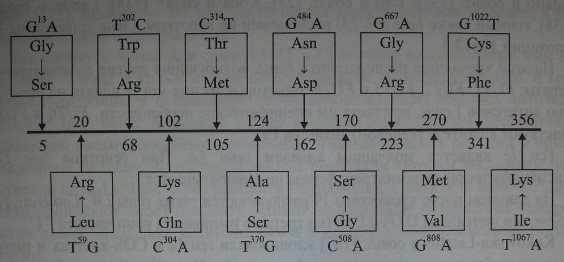
Мутации в гене FUT3 у лиц с фенотипом Lewisnuir Двенадцать нуклеотидных замен, приводящих к продукции неактивной фукозилтрансферазы, по Cooling, Gu [59].
А в нуклеотиде 667. Они приводили к замене Cys 341 Щ Phe, Не 356 Ш Lys, которые также сочетались с низкой активностью фукозилтрансферазы.
Две мутации, Trp 68 —► Arg и Thr 105 —> Met, найдены Elmgren и соавт. [71]. Для того чтобы определить их влияние на активность фукозилтрансферазы, Elmgren и соавт. [70] сконструировали 2 химеричных протеина: FUT3 с Trp 68 —> Arg и FUT3 с Thr 105 —> Met. При первой замене продуцировался фермент с низкой активностью, тогда как при второй замене активность фермента соответствовала норме. Авторы пришли к выводу, что гомозиготность по Trp 68 —* Arg дает фенотип Le(a-b-).
Orntoft и соавт. [187] описали мутацию (С 445 А) у человека Le(a-b-), больного раком. Эта мутация приводила к продукции метионина в позиции 146, однако такая же мутация обнаруживалась и у здоровых лиц.
Mollicone и соавт. [171] установили, что примерно 9 % жителей острова Ява не имеют <х(1,3)-фукозилтрансферазы (FUT6), обычно присутствующей в плазме человека, однако содержат антигены Lewis.
При клонировании гена FUT5 от лиц с дефицитом а(1,3)-фукозилтрансферазы найдены 3 точки мутаций: Arg 173 Щ Cys, Pro 187 Ш Leu, Thr 388 —► Met, которые, однако, не сказывались на активности фермента.
При клонировании гена FUT6 выявлены замены Pro 124 —► Ser, Glu 247 —> Lys, Туг 315 —► стоп-кодон. Две последние замены приводили к продукции неактивного фермента. Замены Glu 247 т Lys и Tur 315—» стоп-кодон в FUT6, которые сочетались с дефицитом а(1,3)-фукозилтрансферазы, обнаружены среди полинезийцев и шведов (Larson и соавт. цит. по [115]). Таким образом, ген FUT6 отвечает за активность <х(1,3)-фукозилтрансферазы в плазме у людей и как другие гены FUT полиморфен.
- Категория: Системы Lutheran и Lewis
Ингибиция Lewis-антител
Lewis-антитела нейтрализуют с целью идентификации антител другой специфичности, присутствующих в сыворотке. Для этого используют образцы слюны, тестированные на наличие субстанций Lea, Leb и АВН, а также вытяжки из гуммиарабика или синтетические антигены Lewis (Spitalnik и соавт., цит. по [115]). Цельная слюна нередко вызывает гемолиз эритроцитов, поэтому ее предварительно разводят раствором натрия хлорида и кипятят, чтобы устранить присутствующую в ней слизь. Методика подробно описана П.Н. Косяковым [6] и Judd [125].
Mollison и соавт. [173] описали случай, когда пациенту Le(a-b-), имеющему антитела анти-Le3 + Leb, гемолизирующие эритроциты в тестах in vitro, с целью нейтрализации антител ввели парентерально очищенный концентрат Le3 и Leb. Антитела пациента были нейтрализованы in vivo, после чего ему было перелито несколько доз крови Le(a-b+) без каких-либо реакций. Через несколько дней антитела анти-Le3 и aHra-Leb снова появлялись в плазме пациента, теперь уже в более сильной форме, чем до нейтрализации. Прямая проба Кумбса стала положительной, поскольку перелитые ранее эритроциты были Le(a-b+) и сенсибилизировались антителами in vivo, однако гемолиза не наблюдалось. Вскоре циркулирующие эритроциты приобрели фенотип реципиента Le(a-b-) и прямая проба Кумбса стала отрицательной.
Нейтрализация антител in vivo была применена также Andorka и соавт., Pelosi и соавт. (цит. по Issitt, Anstee [115]), но широкого распространения не получила: в значительной степени в связи с тем, что в этих модельных экспериментах даже сильные антитела, деструктирующие эритроциты in vitro, не проявляли реактогенных свойств in vivo.
Хромосомная локализация
По данным Mollicone и соавт. [172], Nishihara и соавт. [179], Kukowska-Latallo и соавт. [137], Ball и соавт. [28], Koda и соавт. [134], Lamm и соавт. [138], генный локус Lele (FUT3) расположен на коротком плече хромосомы 19 в позиции 19р13.3.
На этой хромосоме располагаются гены, кодирующие другие фукозилтранс-феразы: FUT1, FUT2, FUT5 и FUT6. Некоторые из них могут влиять на продукцию антигенов Lewis, придавая им специфические особенности. FUT6 и FUT2 участвуют в синтезе тканевого антигена Ley.
Ген к является молчащим аллелем гена Le. При генотипе lele Le-генспецифическая фукозилтрансфераза не вырабатывается.
На длинном плече хромосомы 19 располагаются также гены LW, Lutheran, Eh и Sese, ген пептидазы D {Pep D) и ген третьего компонента комплемента - С,.
Kukowska-Latallo и соавт. [137] клонировали ген Le в COS-клетках и получили рекомбинантные фукозилтрансферазы <х(1—»3) и а(1—>4), способные синтезировать антигены Lea, Leb и X (SSEA-1). Клонированные трансферазы являлись продуктом именно Ze-гена и были однотипны с Lewis-трансферазой, полученной из человеческого молока [151,195].
Suh и соавт. [220] описали несколько мутаций в генах FUT2 и FUT3, при которых антигены Lewis не вырабатывались, f'
- Категория: Системы Lutheran и Lewis
Большинство антител Lewis не способно разрушать эритроциты, содержащие соответствующие антигены, in vivo [111]. Холодовые антитела анти-Le IgM, температурный оптимум которых ниже комнатной температуры 20-22 °С, не активны при температуре тела человека. Неполные антитела анти-Le IgG, хотя и являются тепловыми, однако не вызывают посттрансфузионных осложнений как истинные антиэритроцитарные антитела.
Выделяют четыре причины, по которым Lewis-несовместимость не проявляет себя клинически [115].
Во-первых, большинство антител, особенно анти-Le15, имеет низкую активность. Многие из них не реагируют с нативными эритроцитами и определяются только с помощью стандартных эритроцитов, обработанных протеолитически-ми ферментами (фицин, папайи, бромелин). Такие ферментзависимые антитела в реакциях несовместимости in vivo не участвуют.
Во-вторых, антигены Lewis, присутствующие в донорской плазме (если переливают цельную кровь) и в том небольшом количестве плазмы, которое имеется в эритроцитарном концентрате, нейтрализуют антитела Lewis реципиента. Взаимодействие антител с растворенными в плазме субстанциями происходит быстрее, чем с субстанциями, фиксированными на мембране эритроцитов, и деструкция эритроцитов не успевает развиться.
В-третьих, перелитые эритроциты утрачивают свои антигены Lewis в плазме реципиента. Например: эритроциты Le(a-b+), перелитые реципиенту Le(a-b-), имеющему антитела анти-Le15, становятся Le(a-b-), а олигосахариды Leb, смытые с эритроцитов, нейтрализуются присутствующими антителами анти-Le15.
В-четвертых, если антитела имеют специфичность aHTH-LebH, они способны реагировать только с эритроцитами OLe(a-b+), но не эритроцитами ALe(a-b+). Таким образом, для реципиентов группы А(П), содержащих антитела aHTH-LebH, любые эритроциты А(П) будут совместимыми.
Совместимость донора и реципиента по групповым антигенам, растворенным в плазме крови, в трансфузиологии не учитывают.
Иммуносерологам и трансфузиологам известно, что универсальную плазму группы AB(IV), содержащую группоспецифические субстанции АВО, переливают в большом количестве (2-2,5 л и более) реципиентам А(И) и В(Ш), имеющим естественные антитела против этих антигенов. Однако при этом каких-либо осложнений не наблюдают.
Антигены системы Gm при переливании плазмы также не учитывают, хотя некоторые реципиенты (по нашим подсчетам, около 5 %) содержат антитела против иммуноглобулинов переливаемой плазмы. Однако и в этих случаях трансфузии не сопровождаются реакциями.
По-видимому, взаимодействие растворимых антигенов с антителами не приводит к посттрансфузионным реакциям, но когда мишенью становятся нерастворимые антигены, фиксированные на эритроцитах, последние разрушаются, что проявляется клинически в виде тяжелого посттрансфузионного осложнения.
Антигены Lewis, являясь по своей природе водорастворимыми, в основной своей массе находятся в плазме и реагируют с соответствующими антителами в жидкофазной системе. Эритроциты, содержащие антигены Lewis в значительно меньшем количестве, чем плазма, остаются интактными. Это, на наш взгляд, пятая причина того, почему несовместимость донора и реципиента по системе Lewis не вызывает посттрансфузионных реакций.
В экспериментах по приживлению эритроцитов, меченных Cr51, in vivo установлено, что время циркуляции эритроцитов Le(a~b+) у реципиентов, имеющих анти-Ееь-антитела, такое же, как у реципиентов без антител. Немедленных или отсроченных трансфузионных реакций не наблюдали. Уровень гемоглобина после переливания Ееь-несовместимых эритроцитов повышался как при совместимой трансфузии и оставался одинаковым в течение нескольких недель.
При огромном объеме трансфузиологической помощи в современной медицинской практике лрансфузионные реакции, вызванные антителами анти-Le3, все-таки регистрируют (М.А. Умнова и др. [12], Krieger, Simmons [136], Brendemoen, Aas [43] и др. [160,164,191]).
Jesse, Sheek [119] описали острую гемолитическую реакцию средней тяжести у 21-летней африканки, которой в связи с осложненным спонтанным выкидышем 4 раза перелили кровь. Перекрестные пробы на индивидуальную совместимость (тест с центрифугированием и непрямая реакция Кумбса) перед каждой трансфузией были отрицательными. После 4-го переливания у женщины появились боли в пояснице, озноб, температура, гематурия. Кроме анти-Ееь-антител, в сыворотке женщины других антиэритроцитарных антител не обнаружено. Гемолитическая реакция была кратковременной, осложнение через 2 дня было купировано. Авторы полагали, что картина посттрансфузионного осложнения была вызвана анти-Ееь-антителами. Другие случаи гемолитических посттрансфузионных реакций, обусловленных антителами анти-Leb, не описаны.
Aubuchon и соавт. [26], van Loghem и соавт. (цит. по [6]) привели случаи посттрансфузионных реакций, обусловленных антителами анти-Leх. Для таких реципиентов практически все доноры несовместимы. Однако даже в этих случаях, как отмечают Waheed и соавт. [233], посттрансфузионные реакции крайне редки.
В акушерской практике зарегистрированы единичные случаи гемолитической болезни новорожденных, связанные с Lewis, которые, однако, вызывали сомнение относительно их обусловленности именно этим фактором. Mollison и
соавт. [173], анализируя эти работы, не нашли достаточного клинического и гематологического подтверждения.
Как показали Spitalnik и соавт. [213], в 12 случаях из отобранных ими 13 пар мать - плод в крови матери присутствовали одновременно IgM и IgG анти-Ьеа-антитела, в пуповинной крови новорожденных имелись антитела анти-Le8 IgG, регистрируемые только высокочувствительным ферментсвя-зывающим имммуносорбентным методом. Гемолитических реакций у новорожденных не бвло. Полученные данные позволили авторам заключить, что антитела анти-Le3 IgM через плаценту не проходят, а антитела анти-Leа IgG легко проникают через плаценту, однако не вызывают разрушения эритроцитов плода, поскольку антигены Lewis у плодов и новорожденных отсутствуют.
- Категория: Системы Lutheran и Lewis
AHTH-AjLe5 и анти-ВЬеь
В 1968 г. Seaman и соавт. [206] при выполнении пробы на индивидуальную совместимость эритроцитов донора с сывороткой больного Siedler нашли антитела анти-А^е13. Сыворотка реагировала с эритроцитами, содержащими оба антигена - Ах и Leb, но не реагировала с эритроцитами OLe(b+) и AjLe(b-), содержащими эти антигены порознь. Вскоре 2 такие же сыворотки были найдены Grookston и соавт. [61] и Gundolf [94].
Антитела анти-А^еь нейтрализуются слюной всех секреторов А независимо от Lewis-групповой принадлежности (Grookston и соавт. [61]).
У лиц с фенотипом A1Leb антиген AjLeb присутствует в слюне.
Наряду с антителами анти-А^е* встречаются антитела aHTH-BLeb, реагирующие с антигеном Leb, когда тот присутствует на эритроцитах вместе с групповым антигеном В.
Антитела анти-А^е13 и aHTH-BLeb направлены против олигосахаридов Lebтипа 1, к которым добавлены имммунодоминантные сахара А или В, что и приводит к формированию антигенных детерминант, определяемых указанными антителами.
Для сравнения: антитела aHTH-LebL реагируют только с эритроцитами 0(1), A2Le(b+); а анти-А Leb и aHTH-BLeb - с эритроцитами, несущими Leb и соответствующую А- или В-детерминанту.
Tilley и соавт. [228], Crookston и соавт. [90] установили, что у людей AtLebи BLeb в плазме присутствуют гликосфинголипиды, несущие соответственно антигены AjLeb и BLeb. Такие лица по набору генов относятся к A1, Le, Se и В, Le, Se соответственно. Насколько известно, ни анти-А^еь, ни aHTH-BLeb не дают реакций при переливании компонентов крови (Issitt, Anstee [115]).
Анти-А^е11 и анти-ВЬей
В 1958 г. Andresen [14], обследуя больного раком желудка с фенотипом A2Le(a~b+), нашел антитела, которые реагировали с эритроцитами AjLe(a-b-) секреторов, менее сильно - с эритроцитами АХе(а-Ь-) секреторов. Автор предположил, что ген Se в отсутствие гена£е специфически видоизменяет экспрессию антигена А и что обнаруженные им антитела (известные в литературе как Magard-антитела) выявляли антиген A1Led. Последующие исследования (Hirsch и соавт., 1975) подтвердили предположение Andresen. Magard-сыворотка была первой из обнаруженных сывороток со специфичностью анти-А1Ьес1. Эти антитела, как теперь известно, реагируют с олигосахаридами типа 1А (производными цепей типа 1Н). Анти-А1Ьес1, так же как aHTH-BLed, не дают трансфузионных реакций.
- Категория: Системы Lutheran и Lewis
Анти-Ьеа
Следует подчеркнуть, что антитела анти-Ьеа реагируют одинаково активно с эритроцитами Le(a+) независимо от того, к какой группе по системе АВО последние относятся. В противоположность этому антитела анти-Ьеь не столь однотипны. Некоторые из них реагируют предпочтительно с эритроцитами Le(b+) группы 0(1) и А2(П), другие - с эритроцитами Le(b+) группы А^П), В(Ш) и A1B(IV).
Miller и соавт. [166] наблюдали, что слюна доноров, имеющих антитела анти-Ьеа, содержит субстанции А, В и Н, но лишена субстанций Lea и Leb.
Некоторые сыворотки антаьЬеа гемолизируют эритроциты Le(a+) в присутствии комплемента и могут быть использованы в реакции связывания комплемента.
Обработка эритроцитов протеолитическими ферментами усиливает реактивность сывороток Lewis, в том числа анти-Ьеа.
Преципитины анти-Le3, реагирующие со слюной лиц Le(a+), несекрети-рующих АВН, описаны Ueyama [229, 230], Furuhata и Ueyama [80]. Антитела были найдены в сыворотках кур, интактных и после инъекций слюны несекреторов 0(1) группы.
Преципитирующие и агглютинирующие сыворотки анти-Ьеа получены от кур [267, 80], кроликов [47, 114, 145] и коз [130, 156, 158] иммунизацией слюной несекреторов, содержимым овариальных кист несекреторов [27, 47, 114] или эритроцитами кролика, нагруженными субстанцией Lea сыворотки человека [145 ].
Анти-Ьеь^
Как и анти-Ьеа, большинство встречающихся антител анти-Leb относится к IgM неиммунного происхождения. Они являются комплементсвязывающими и нередко их выявляют как слабые сопутствующие антитела в сыворотках, содержащих сильные антитела анти-Ьеа.
В большинстве случаев антитела анти-Ьеь находят у людей группы Afll) или A1B(IV), реже - у людей 0(1) и В(Ш) (Kissmeyer-Nielsen [132]), что связано с неодинаковым распределением на эритроцитах этих лиц вещества Н. В свою очередь сыворотки анти-Ьеь часто содержат некоторое количество антител анти-Н. В силу структурного сходства антигенов Leb и Н эти антитела реагируют особенно сильно с эритроцитами O(I) и А2(П), а слюна, нейтрализующая антитела анти-Ьеь, также ингибирует антитела анти-Н.
Описаны 2 формы антител анти-Ьеь: LebH и LebL [41, 53, 54, 210].
Анти-Ьеьн
Антитела анти-Ьеьн реагируют с эритроцитами Le(b+) 0(1) и А, но не Le(b+) А,(И) и В(Ш). Антитела анти-Ьеьн нейтрализуются слюной людей Le(a-b+) и, что примечательно, слюной людей Le(a-b-) секреторов АВН. Иными словами, анти-Ьеьн могут быть нейтрализованы слюной, которая содержит субстанцию Н или Leb + Н, отсюда название - анти-Ьеьн. 'щ(
Антитела анти-Ьеьн направлены преимущественно к олигосахаридам типа 1 Н, которые присутствуют на эритроцитах 0(1) и А2(П).
На эритроцитах А,(П), В(Ш) и A1B(TV) олигосахаридные цепи типа 1Н закрыты иммунодоминантными сахарами А или В, поэтому недоступны для антител анти-Ьеьн.
Анти-Ьеьн не агглютинируют эритроциты Le(a-b-) АВН-секреторов, которые несут олигосахариды типа 1 Н, т. е. антигены Led, с которыми анти-Ьеьн не связываются, однако ингибируются слюной Le(a-b-) секреторов, как упоминалось вьппе. P*Bird [32, 33] высказал предположение, что антитела анти-Ьеьн могут быть представлены смесью aHTH-Leb+H15 по аналогии с антителами анти-A+Aj и анти-Н+Нг Kissmeyer-Nielsen (цит. по [197]) нашел 2 сильные сыворотки анти-Н у лиц Le(a+b~), которые в подтверждение этой идеи реагировали и с эритроцитами 0(1) Le(a+b-), и с эритроцитами 0(1) Le(a-b-). Wiener и соавт. [235] также считают, что антитела анти-Hj близки по специфичности к анти-Ьеьн.
Kornstad [135] нашел, что антитела лиц Le(a+b-) имеют больший аффинитет к антигену Н, чем к Leb, а антитела лиц Le(a-b-) - больший аффинитет к Leb, чем к Н. На этом основании им высказано предположение о существовании широкой палитры антител: от чистых анти-Н через промежуточные формы анти-H(Leb) и анти-Ьеь(Н) до чистых анти-Ьеь.
Анти-1Ьеьн
Tegoli и соавт., 1971 [225] нашли в сыворотке человека AjLe(a-b-) антитела анти-1Ьеьн, которые реагировали с эритроцитами Le(a-b+) группы 0(1) или А2(П) только в том случае, если эритроциты были I+, что и послужило поводом обозначить их специфичность как анти-1Ьеьн. Второй случай анти-1Ьеьн описали Branch и Powers в 1979 г. (Issitt, Anstee [115 J).
Антитела анти-1Ьеьн нейтрализуются слюной лиц Le(a-b+) группы 0(1) и А, (И) независимо от присутствия у них антигенов Ii (I+, I-i+), но не нейтрализуются слюной лиц Le(a+b+)I+ группы 0(1) и О, (Бомбей).
Анти-Ьеьь
Антитела анти-Ьеьь в противоположность анти-Ьеьн реагируют с эритроцитами Le(b+) независимо от их групповой принадлежности по системе АВО. От анти-Ьеьн они отличаются тем, что нейтрализуются слюной, содержащей субстанции Leb+H, но не ингибируются слюной, содержащей субстанцию Н без Leb.
Антитела анти-Ьеьь направлены к L-фукозным остаткам олигосахаридных цепей типа 1, которые в процессе синтеза на эритроцитах А^П), В(Ш) и A^IV) не закрываются иммунодоминантными сахарами А и В.
Как указывали Race и Sanger [197], в первые годы исследования системы Lewis данные о частоте антигена Leb одних авторов не соответствовали данным других авторов. Разделение антител анти-Ьеь на LebH и LebL внесло ясность в существующее положение. При типировании эритроцитов группы А^П), В(Ш) и AjB(IV) по антигену Leb следует использовать сыворотки anra-LebL, поскольку сыворотки анти-Ьеьн в этом случае будут давать ложноотрицательные результаты.
Частота анти-Ьеа и анти-Ьеь
Антитела Lewis чаще выявляют методом солевой агглютинации на плоскости или в пробирках при комнатной температуре. В методиках, предусматривающих инкубацию ингредиентов реакции при 37 °С, антитела Lewis обнаруживают с меньшей частотой, за исключением проб, в которые для усиления реакции добавляют полиэтиленгликоль, полибрен или 10% желатин. В этой среде Lewis-антитела IgM и IgG проявляют одинаково высокую активность.
Сильные сыворотки анти-Le3 находят редко, а обычно встречающиеся имеют достаточно высокую частоту.
По данным Salmon и соавт. (цит. по [115]), среди 72 ООО парижан обнаружено 249 лиц, содержащих антитела анти-Le3, 31 - anra-Leb, 49 - airra-Lex. Из 31 сыворотки anra-Leb 29 были airoi-LebH, 2 - anra-LebL. Частота антител составила 0,45 %.
Среди 40 ООО обследованных пациентов клиник Дюк Университета Issitt и Anstee [115] выявили 167 человек с антителами анти-Le3 и 39 - с aHTH-Leb. Частота антител составила 0,49 %. Как отмечают авторы, антитела Lewis встречаются в 3-4 раза чаще у негров, чем у белых, поскольку среди негроидов лица с фенотипом Le(a-b-), являющиеся основными продуцентами антител, составляют 20-30 %, а среди европеоидов - только 6 %.
Методы исследования антител Lewis также влияют на частоту их обнаружения. С использованием эритроцитов, обработанных ферментами, антитела Lewis выявляют существенно чаще.
Анти-Ьес
Антитела aHra-Lec находят в сыворотках Le(a-b+) секреторов. Эти антитела нейтрализуются слюной Le(a-b-) несекреторов, менее сильно - слюной Le(a-b-) секреторов и Le(a+b-) несекреторов и очень слабо нейтрализуется слюной Le(a-b+) секреторов. Реакции ингибируются также гликопротеинами, выделенными из содержимого овариальных кист Le(a-b-) несекреторов и трисахаридом фукозиллактозы. Другие сахара не обладают способностью нейтрализовать антитела aHTH-Lec.
Антитела anra-Lec найдены Gunson и Latham [95] у людей, однако более сильные реактивы получают иммунизацией животных [7, 86, 109, 114].
Iseki, Masakia и Shibasaki [114] описали антитела, названные ими анти-Ьес,которые были получены путем иммунизации кроликов слюной Le(a-b-) секреторов и адсорбцией иммунных сывороток энзимированными эритроцитами Le(a+b-) и Le(a-b+). Результат исследования 485 японцев сыворотками анти-Le3, анти-Le15 и полученными сыворотками анти-Ьес был следующим (по Race, Sanger [197]): 107 (22,06 %) - Le(a+b-c+), 396 (76,08 %) - Le(a-b+c-), 9 (l,86%)-Le(ab-c+).
М.И.£5 Потапов [9] получил антитела анти-Le иммунизацией коз слюной лиц Le(a-b-) несекреторов. Полученные иммунные сыворотки были адсорбированы трипсинизированными эритроцитами Le(a+), Le(d+) и Le(b+). Оставшиеся антитела реагировали с эритроцитами 0(I)Le(a-b-c+), но не 0(1) Le(a-b-c-d+).
Race и Sanger [197] сравнили специфичность козьей сыворотки, полученной М.И. Потаповым, и человеческой сыворотки анти-Ьес, найденной Gunson и Latham [95]. Результаты исследования полностью совпали: эритроциты Le(a-b+) несекреторов агглютинировались обеими сыворотками, эритроциты Le(a-b-) секреторов не агглютинировались.
Анти-Ьей
Эти антитела реагируют с эритроцитами Le(a-b-c-) лиц leSe/leSe и lese/leSe. Они направлены против олигосахаридов типа 1 Н, которые, так же как и другие олигосахариды Lewis, адсорбируются на эритроцитах из плазмы [109].
Антитела aHTH-Led получают иммунизацией животных [7, 10, 109] слюной выделителей АВН. Аллогенные антитела aHTH-Led в литературе не описаны.
Анти-Le* (анти-ЬеаЬ)
У лиц Le(a-b-) находят антитела, которые по своей направленности ведут себя как aHTH-Lea + анти-Le15. Они могут быть простой смесью aHTH-Lea и анти-Leb. В этом случае фракция aHTH-Lea легко нейтрализуется добавлением слюны донора Le(a+b-), в которой имеется субстанция Lea, но нет субстанции Leb. Адсорбированная таким образом сыворотка содержит только антитела анти-Le15. Слюну лиц Le(a-b+) для дифференциальной нейтрализации не применяют, поскольку она содержит субстанции Lea и Leb и полностью истощает оба антитела. Антитела aHTH-Lea можно удалить также дифференциальной адсорбцией эритроцитами Le(a+b-).
Если антитела анти-Le3 и анти-Le15 представляют собой не смесь, а одно связанное антитело анти-Le315, обозначенное анти-Le* [15, 122, 123], сепарацию антител дифференциальной нейтрализацией или адсорбцией провести не удается.
Sturgeon и Arcilla [218] отметили, что антитела анти-Leх реагируют с эритроцитами Le(a+b~), Le(a-b+) и эритроцитами новорожденных, имеющих, как известно, фенотип Le(a-b-).
Тот факт, что эритроциты детей Le(a-b-) не реагируют с моновалентными воротками I и анти-Ьеь, но реагируют с анти-Ьех, свидетельствует о существовании на эритроцитах детей антигена Le\
Антиген Lex присутствует также у некоторых взрослых.
Таким образом, антитела анти-ЬеаЬ (анти-Ьех) не являются простой смесью антител и выявляют, помимо антигенов Lea и Leb, третий специфический антиген системы Lewis - Lex (Leab).
Andresen [16] первоначально полагал, что продукция Lex зависит от дополнительного гена Lex в системе Lewis, однако более поздние исследования, проведенные Sturgeon и Arcilla [218], позволили заключить, что продукция антигенов Lea, Leb и Lex является результатом действия одного гена Le.
В работах некоторых авторов высказываются сомнения относительно того, что Lex такой же самостоятельный антиген, как Lea и Leb, поскольку по серологической характеристике антитела aHTH-Lex представляют собой комбинацию специфичности aHTH-Lea + aHTH-Leb, подобно перекрестно реагирующим антителам ар (анти-С) системы АВО.
Реакции антител aHTH-Lex с эритроцитами Le(a-b-) новорожденных были объяснены тем, что многие образцы пуповинной крови реагируют с сильными сыворотками aHTH-Lea в непрямой пробе Кумбса [62]. Иными словами, эритроциты новорожденных содержат некоторое количество вещества Lea, с которым реагируют антитела amn-Lex.
Антитела aHTH-Lea обычно вырабатываются секретерами субстанций АВН, антитела анти-Le15 - несекреторами. Антитела aHTH-Lex часто, но не всегда вырабатываются Le(a-b-) секреторами. В этом их сходство с антителами aHTH-Lea.
Одни лица продуцируют антитела aHTH-Lea или aHTH-Leb, другие могут продуцировать оба антитела в виде раздельных фракций (aHTH-Lea + анти-Leb) или одной фракции aHTH-Leab (aHTH-Lex).
Arcilla и Sturgeon [21-23] показали, что амниотическая жидкость содержит высокий уровень субстанции Lea, которая проявляет себя серологически как Lex.
Антитела aHTH-Lex нейтрализуются слюной, содержащей Le3, слабее - слюной Leb и, что удивительно, слюной лиц Le(a-b-), являющихся несекреторами [16, 19]. Ингибиция слюной Leb (т. е. слюной секреторов) более сильная, чем слюной лиц Le(a-b-) несекреторов. Уместно напомнить, что лица Le(a-b-) могут быть секреторами и несекреторами.
То обстоятельство, что антитела aHTri-Lex ингибируются слюной Le(a-b-) несекреторов, подтверждает существование антигена Lex как самостоятельной единицы.
Химическая структура антигена Lex, как полагают Schenkel-Brunner и соавт. [203, 205] и другие авторы [48,106, 185, 240], близка детерминантам, определяемым с помощью антител анти-Le3 и aHra-Leb.
Для сравнения: перекрестно реагирующий антиген С в системе АВО (по Винеру) присутствует на эритроцитах А и В. Антитела анти-С представляют собой несепарируемый агглютинин ар (анти-А,В), присутствующий в сыворотках лиц 0(1) наряду с сепарируемыми аир.
В системе резус описаны антитела анти-DC, реагирующие с антигенами D, С и антигеном G, который, как правило, сопровождает D и С, но иногда встречается на DC-отрицательных эритроцитах (фенотип cdeG).
Таким же перекрестно реагирующим антигеном, по-видимому, является антиген Lex в системе Lewis.
Антитела affra-Lex (как aHTH-Lea и aHTH-Leb) обычно имеют не аллоиммун-ное происхождение, обладают способностью связывать комплемент и могут проявлять гемолитические свойства in vitro.
- Категория: Системы Lutheran и Lewis
Антитела Lewis (aHTH-Lea, aHTH-Leb, анти-Le* и другие) встречаются, как правило, у людей с фенотипом Le(a-b-) [92, 166] и редко имеют трансфузионное происхождение. Их появление может совпасть с переливанием компонентов крови, но обусловлено другими причинами.
Антитела анти^еа-специфичности встречаются чаще других (Jordal [122, 123], Kissmeyer-Nielsen [132]).
Мы наблюдали 2 случая антител Lewis у молодых мужчин [5]. Оба донора военнослужащие, 20 и 22 лет. У одного из них, A(II) CcDEe kk Le(a-b-), в сыворотке при определении группы крови перекрестным методом выявлены холодовые агглютинины с титром 1 : 128, реагирующие на плоскости. Обнаруженные антитела агглютинировали эритроциты Le(a+b~), не реагировали с собственными эритроцитами и эритроцитами доноров Le(a-b+) и были идентифицированы как анти-Ьеа.
От 2 донаций крови получили 350 мл сыворотки, которая в разведении не менее чем в 2-3 раза продолжительное время служила как высокоактивный тестовой реактив анти-Ьеа. Интересная деталь: в порции сыворотки от третьей до-нации (через год после первой) антитела анти-Ьеа отсутствовали.
Issitt и Anstee [115] (см. выше) привел случай неожиданного исчезновения антител анти-Ьеа у женщины вскоре после родов.
Другой донор, AB(IV) CcDee kk Le(a-b-), содержал антитела, которые реагировали с 7 образцами эритроцитов Le(a+b-), 30 образцами Le(a-b+), но не реагировали с собственными эритроцитами и эритроцитами донора Le(a-b-) и по своей специфичности относились к анти-ЬеаЬ(Ьех). Особенностью этих антител являлось то, что они агглютинировали эритроциты, обработанные проте-олитическими ферментами (проназой С), но не реагировали с нативными эритроцитами, подобно сыворотке первого донора.
Оба донора не имели в анамнезе инъекций или трансфузий каких-либо компонентов крови.
Антитела Lewis описаны в литературе в основном как «naturally occurring)) -спонтанные, естественно встречающиеся. Относительно их природы нет единого мнения. Некоторые авторы полагают, что они вырабатываются в результате контакта с Lewis-олигосахаридами, распространенными в окружающей среде.
По нашему мнению, Lewis-антитела имеют такое же естественное происхождение, как изогемагглютинины АВО. Они компенсируют отсутствие Lewis-олигосахаридов в организме людей Le(a-b-) так же, как изогемагглютининь1 а и Р компенсируют отсутствие полисахаридов А и В у лиц О, А и В. Однако Lewis-антитела в отличие от групповых изогемагтлютининов имеются не у всех людей.
Следует подчеркнуть, что естественные Lewis-антитела встречаются у европеоидов с весьма высокой частотой - 0,49 % (Issitt, Anstee [115]). У людей Le(a-b-) частота Lewis-антител, по нашим расчетам, составляет 5,8 %, что не может иметь характер случайного явления. Среди негроидов, у которых частота лиц Le(a-b-) достигает 20-22 %, Lewis-антитела встречаются в 3-4 раза чаще. На наш взгляд, это подтверждает суждение о том, что система Lewis построена по такому же принципу, как АВО: есть антигены - нет антител, нет антигенов - есть антитела.
Находят антитела и иммунного происхождения, в том числе появившиеся в результате трансфузий и беременностей.
Не исключено, что большинство Lewis-антител не определяются существующими методами и их реальная частота значительно выше выявляемой. Система Lewis, как неоднократно подчеркивалось, отличается своеобразием.
Обычно антитела анти-Le3 находят при проведении пробы на индивидуальную совместимость перед переливанием эритроцитов или при определении группы крови перекрестным методом. По данным Mollison, Engelfriet, Contreras [173], анти^еа-антитела - полные холодовые агглютинины IgM, хорошо реагируют на плоскости при комнатной температуре. Встречаются также неполные тепловые IgG анти-Ьеа-антитела [173].
Holburn [ПО] отметил, что антитела анти-Le3, содержащиеся в сыворотке, нередко представлены комбинацией иммуноглобулинов IgM и IgG одинаковой специфичности. Комбинированные антитела проявляют более высокую активность по сравнению с антителами, встречающимися по отдельности.
Антитела анти-Le3 IgM имеют более высокую скорость связывания с антигеном и сильнее активируют комплемент, чем IgG. Сводные данные о свойствах Lewis-антител приведены в табл. 9.8.
Таблица 9.8
Характеристика Lewis-антител
|
Показатели |
Реакция антител |
|||||||
|
Lea |
LebH |
LebL |
Lex |
Lec |
Led |
AjLeb |
||
|
Реакция |
в солевой среде |
+ |
+ |
+ |
| |
1 |
± |
+ |
|
с антиглобулином |
+ |
+ |
+ |
|
+ |
± |
+ |
|
|
с ферментами |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Класс иммуноглобулинов |
М, +G |
М |
М |
M,+G |
G? |
G? |
| . М |
|
|
Связывание комплемента |
+ |
| |
+ |
|
|
|
+ |
|
|
Гемолиз in vitro |
+ |
— |
1 |
|
|
|
+ |
|
|
Посттрансфузионные реакции |
Редко |
Нет |
Нет |
Редко |
Нет |
Нет |
Нет |
|
|
Гемолитическая болезнь новорожденных |
Нет |
Нет |
Нет |
Нет |
Нет |
Нет |
Нет |
|
|
Частота реагирования, % ** |
22 |
*** |
72 |
94 |
1 |
6 |
**** |
|
* с эритроцитами Le(a+) гемолиз более выражен, чем с эритроцитами Le(a-); ** среди европеоидов; *** сыворотки анти-Leьн реагируют только с эритроцитами Le(b+) группы O(I) и А2 (II); **** сыворотки анти-А^еь реагируют только с эритроцитами Le(b+) группы А(П).
Как упоминалось выше, почти все антитела анти-Le3 вырабатываются у людей Le(a-b-). Лица Le(a-b+) секретируют оба вещества (Le3 и Leb), поэтому их сенсибилизации к антигену Le3 не происходит. Однако, как показали Judd и соавт. [126], Cowles и соавт. [60], Issitt, Anstee [115], в редких случаях у лиц Le(a-b+) антитела анти-Le3 могут присутствовать.
В одном из случаев (Judd и соавт. [126]) антитела анти-Le3 были обнаружены у больного карциномой пищевода и, хотя его слюна содержала субстанции Н, Le3 и Leb, количество антигена Leb на его эритроцитах было в 2 раза меньше, чем на эритроцитах Le(a-b+) здорового человека. Авторы полагают, что онкогенный статус пациента настолько блокировал синтез антигенов Lewis, что организм больного стал распознавать антиген Le3 как чужеродный. Авторы высказали предположение, что антителообразование в рассматриваемом случае могло иметь также аутоиммунную природу, поскольку эти антитела, хотя и не реагировали с эритроцитами пациента, но полностью ингибировались его слюной. В связи с тем, что они были инертны по отношению к собственным эритроцитам, сокращения продолжительности жизни эритроцитов in vivo не наблюдали. Антитела имели доброкачественный неаутоагрессивный характер.
Miller и соавт. [166] полагают, что на способность лиц Le(a-b-) продуцировать анти-Ьеа-антитела влияет их секреторный статус. По наблюдениям этих авторов, антитела анти-Ьеа вырабатывались лицами Le(a-b-) выделителями АВН, т. е. имеющими комбинацию генов le и Se. Issitt и Anstee [115] усматривают в этом противоречие. Согласно существующим представлениям при генотипе lese/leSe L-фукоза соединена с терминальным участком олигосахаридной цепи типа 1. Не ясно, почему лица Le(a-b-) выделители (генетически lese/leSe), имеющие L-фукозу на терминальном участке, вырабатывают антитела, распознающие L-фукозу, прикрепленную к рядом расположенному субтерминальному участку.
Группа крови АВО сильнее влияет на продукцию aHTH-Lea. Park и соавт. [188], Kissmeyer-Nielsen [132] установили, что среди носителей антител aHTH-Lea чаще встречаются лица А(П), В(Ш) и AB(IV), реже 0(1) по сравнению с частотой этих групп крови в рандомизированной выборке. Так, среди продуцентов aHTH-Lea всего лишь 11 % лиц 0(1) группы, а при нормальном распределении у европеоидов частота этой группы около 33 %. Это объясняется тем, что люди 0(1) продуцируют большое количество субстанции Н, структурно близкой субстанциям Lewis.
Антитела системы Lewis, как уже упоминалось, вырабатываются лицами Le(a-b-), которые не секретируют субстанции АВН (lese/lese). Лица Le(a-b-), секретирующие вещества АВН, то есть генетически leSe/leSe или, как минимум, lese/leSe, синтезируют олигосахариды типа 1 Н. Напомним, что олигосахариды типа 1 Н представляют собой не что иное как антиген Led, близкий по структуре к Leb. Естественно, что при столь близком антигенном родстве Led и Leb лица Le(a-b-d+) не распознают антиген Leb как чужеродный и не продуцируют соответствующие анти-Leь-антитела (см. рис. 9.1,9.2).
Лица Le(a+b-) способны продуцировать антитела aHTH-Leb, поскольку они генетически Lese/Lese или lese/Lese и не синтезируют вещество Leb. Однако в действительности антитела анти-Le15 у лиц Le(a+b-) встречаются крайне редко [41, 82,135], в основном эти антитела присущи лицам Le(a-b-) невыделителям, генетически lese/lese.
Сыворотки aHTH-Leb часто содержат некоторое количество антител анти-Н и имеют свойства, общие с анти-Н: реагируют особенно сильно с эритроцитами О и А2, а слюна, нейтрализующая aHTH-Leb, ингибирует анти-Н.
Многообразие перекрестных реакций, свойственное антителам Lewis, Н, АВО, их способность связываться с группоспецифическими субстанциями слюны подчеркивают не только большое структурное сходство антигенов этих систем, но и большое их разнообразие.
Большое количество моноклональных антител, полученных с целью серологической диагностики опухолевых клеток различных типов, проявляет специфичность анти-Lewis [13,45, 77, 79, 84,116,127,143,212,216].
Антитела, похожие по серологическим свойствам на анти-Ьеа, присутствуют в некоторых лекарственных травах. Лекгин из семян Griffonia Simplicifolia проявляет специфичность анти-Ьеа [208], а также реагирует с антигеном X и Y(Ley) [128,214].
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}