Поиск по сайту
- Вы здесь:
-
Главная

-
Новости
-
Наш блог
-
book
-
Группы крови человека
- Система Kell
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
Система Kell
Антиген Km фенотипически тесно связан с антигенами Kell и Кх и так же, как антигены системы Kell, денатурируется при обработке эритроцитов АЕТ. На основании этого антиген Km был отнесен к системе Kell и получил обозначение ISBT KEL20. Однако принадлежность его к системе Kell или Кх окончательно не доказана и остается принятой лишь условно. Основанием для сомнений служат следующие обстоятельства. Если бы антиген Km относился к системе Kell, то он должен был присутствовать на эритроцитах McLeod, как и другие Kell-антигены, хотя бы в небольшом количестве. Однако антиген Km на эритроцитах McLeod не обнаруживали ни методом прямой агглютинации с сывороткой анти-Km, ни с помощью метода адсорбции - элюции. Более того, лица с фенотипом McLeod продуцируют анти-Кт-антитела, что еще раз свидетельствует об отсутствии антигена Km на их эритроцитах. Из указанного выше очевидно, что гликопротеин Kell не является носителем антигена Km, а KEL и Km - разные гены.
Антиген Km отсутствует на эритроцитах К, то есть не представлен на Кх-протеине и, соответственно, не кодируется геном ХК.
Поскольку антиген Km фенотипически явно ассоциирован с антигенами системы Kell и Кх, данные о том, что он отсутствует на гликопротеине Kell и протеине Кх, не находили объяснения. Issitt и Anstee [204] высказали предположение, что при определенных комбинациях белковых компонентов мембраны, кодируемых независимыми локусами, могут появляться новые антигенные детерминанты, не свойственные этим локусам. Так, продукция Kell-гликопротеина, не содержащего вещества Km, контролируется генами хромосомы 7, а продукция Кх-протеина, также не содержащего вещества Km, контролируется геном хромосомы X. Km-антиген появляется, по-видимому, в результате сочетанного эффекта Kx-KEL в процессе формирования комплекса Кх - Kell.
Не исключено существование самостоятельного гена Km, отсутствующего у лиц с фенотипом К или McLeod, однако какие-либо факты в пользу этого предположения отсутствуют.
Система Кх
Система Кх включает 1 антиген - Кх (ISBT 019001 или ХК1). Redman и соавт. [326] выделили протеин Кх посредством иммунопреципитации анти-Кх-антителами и определили его мол. массу, составляющую около 37 кДа. Затем было установлено, что Кх-протеин фосфорилируется и паль-митилируется [120, 121], но не гликозилируется [198, 224]. Он представлен в мембране эритроцитов 10 трансмембранными доменами [198], состоящими из 444 аминокислот (рис. 5.5).
Предполагаемая топология Кх-протеина в мембране эритроцита представлена на рис. 5.1. Белок Кх связан дисульфидной связью через Cis 347 с Cis 72 гликопротеина Kell. Терминальные участки Кх-протеина, N и С, погружены в клетку.
По мнению Russo и соавт. [331], комплекс Кх - Kell формируется в эн-доплазматическом ретикулуме и транспортируется к клеточной поверхности. Отсутствие вещества Кх в эритроцитах приводит к уменьшению экспрессии Kell-антигенов на поверхности клеточной мембраны (фенотипу McLeod).
Система Кх кодируется Х-сцепленным геном, ХК, и является самостоятельной антигенной системой независимой от Kell, хотя и тесно с ней связанной.
Кх-протеин выражен на эритроцитах лиц с фенотипом Ко сильнее, чем на эритроцитах с обычным Kell-фенотипом [267]. Однако указанная особенность, как полагают Lee и соавт. [238], Carbonnet и соавт. [122], обусловлена не столько количеством Кх-протеина на этих клетках, сколько тем, что в эритроцитах обычного Kell-фенотипа Кх-протеин связан с белком Kell, что мешает ему с той же силой участвовать в реакции агглютинации с сывороткой анти-Кх.
В противоположность антигенам Kell антиген Кх не денатурируется дити-отрейтолом (ДТТ) и 2-аминоэтилизотиоурониумбромидом (АЕТ), которые разрушают дисульфидные связи. Эритроциты, обработанные ДТТ и АЕТ, приобретают серологические свойства эритроцитов Ко с высокой степенью экспрессии Кх-антигена [102,111].
Анти-Кх
У пациента Klaas, имевшего фенотип McLeod (см. Антиген KL), развилась гемолитическая трансфузионная реакция, вызванная антителами анти-KL, которые, как вскоре выяснилось, представляют собой смесь двух антител разной специфичности и могут быть отделены друг от друга с помощью адсорбции и последующей элюции (Van der Hart и соавт. [377], Marsh и соавт. [258, 267]). Антигены, выявляемые с помощью этих антител, получили обозначение Кх и Km, а соответствующие им антитела - анти-Кх и Km.
Анти-Кх-антитела, как установили Marsh и соавт. [267], сильно реагируют с эритроцитами Ко, но не реагируют с эритроцитами McLeod
С эритроцитами обычного Kell-фенотипа они взаимодействуют слабо. Таким образом, антиген Кх присутствует на эритроцитах Ко, отсутствует на эритроцитах McLeod, выражен, хотя и слабо, на эритроцитах лиц, имеющих нормальный Kell-фенотип.
Таблица 5.5
Выраженность антигенов Кх, Km и Kell на эритроцитах лиц с нормальным и Kell-дефицитными фенотипами
|
Фенотип |
Выраженность антигена |
||
|
Кх |
Km |
антигены Kell |
|
|
Обычный |
+ |
++++ |
++++ |
|
McLeod |
— |
— |
+ |
|
К |
++++ |
— |
— |
|
^"mod |
++++ |
Нет данных |
+ |
« + » - антиген слабо выражен;
« ++++ » - антиген сильно выражен;
« - » - антиген отсутствует.
Анти-Кт-антитела, как установили те же авторы [267], реагируют с эритроцитами обычного Kell-фенотипа, но не реагируют с эритроцитами К и McLeod. Считается (Daniels [141]), что антиген Km формируется в том случае, если на поверхности эритроцита экспрессированы оба белка, Кх и Kell. Если один из них редуцирован, антиген Km отсутствует. Похожая ситуация имеет место в системе Rh-Hr (эффект г/иопозиции). Если гены с и Е наследуются в положении цис, помимо антигенов с и Е вырабатывается сопутствующий антиген сЕ. Этот специфический антиген не является смесью антигенов с и Е. Сыворотки анти-сЕ не реагируют ни с антигеном с, ни с антигеном Е в эритроцитах лиц СЕ/се и се/се, но реагируют с антигеном сЕ в эритроцитах лиц cE/cef где с и Е расположены на одной хромосоме. При наследовании указанных генов в положении транс антиген сЕ не вырабатывается.
Антителаити-1^|+ анти-Кт (анти-KL) вырабатываются у лиц с фенотипом McI||o& Анти-Кт-антитела могут вырабатываться также у гомозигот К°/К°, которые не содержат антигена Km. Причиной появления антител являются трансфузии эритроцитов [114, 177, 267, 377], однако известны антитела нетрансфузи-онного происхождения. Carstairs и соавт. [123] привели случай обнаружения таких антител у пациента, не получавшего трансфузий. Антитела у него выработались после перенесенного септического шока.
Антитела анти-Кх и анти-Km могут вырабатываться как вместе, так и по отдельности. White и соавт. [395] описали пациента с McLeod-синдромом, который выработал только анти-Кт-антитела. Кх-антитела в его сыворотке отсутствовали.
Аналогичный случай привели Sharp и соавт. [345]. У больного имелись анти-Кт-антитела, в связи с чем ему потребовался индивидуальный подбор крови. После пяти трансфузий эритроцитов Ко и Kmod, содержащих сильный Кх-антиген, антитела анти-Кх у больного, вопреки ожиданию, не выработались. Авторы отмечают хороший терапевтический эффект трансфузий. Реакцией отсутствовали.
Russo и соавт. [332] нашли анти-Кх-антитела у пациента с McLeod-синдромом, которому в связи с анемией, развившейся вследствие желудочно-кишечного кровотечения, перелили 4 дозы эритроцитов.
Sullivan и соавт. [358] обнаружили анти-Кх-антитела у больного, имевшего обычный Kell-фенотип. Антитела оказались клинически не значимыми и, по-видимому, имели аутоиммунную природу. Они не гемолизировали ни собственные эритроциты пациента, ни перелитые ему эритроциты, содержащие антиген Кх.
Возможная роль антител анти-Кх и анти-Km в развитии ГБН остается для исследователей невыясненной, поскольку эти антитела у женщин не встречаются, а содержатся только у мужчин.

К13-эффект
К13-эффект правильнее называть эффектом отсутствия гена К13, поскольку гаплотип К13-, а не гаплотип К13, содержит ген, подавляющий экспрессию других антигенов системы Kell и napa-Kell (см. К13).
Marsh и соавт. [262], впервые обнаружившие человека с фенотипом К13-, вначале полагали, что низкая экспрессия антигенов k, Kpb, Jsb, Ku и К12 на эритроцитах этого человека связана с его гетерозиготностью по гену К°. Эритроциты пробанда сильно реагировали с сывороткой анти-Кх, что свидетельствовало о наличии у этого человека гаплотипа К°. Генотип пробанда соответствовал формуле К13-/К13-К. Однако, как показали Kaita и соавт. [220], гетерозиготность по гену К° не могла быть причиной депрессии других Kell-антигенов, поскольку люди, являющиеся генетически K°/kKpbJsb, нормально экспрессируют антигены k, Крь и Jsb. Таким образом, Marsh и соавт. [262] оставалось сделать единственно правильный вывод относительно ослабления антигенов Kell у лиц К13-, а именно: низкая экспрессия Kell у лиц К13- обусловлена антитетичным партнером К13, который, будучи в гомозиготной форме и в позиции цис к другим генам KEL, кодирует более слабую, чем в норме, экспрессию антигенов Kell.
У людей К13- слабо выражены антигены К18 (Barrasso [98]), К19 и TOU (Sabo и соавт. [337]).
Вполне вероятно, что случаи ослабления Kell-антигенов, наблюдаемые многими авторами, связаны с гетерозиготностью по К13-, однако это пока невозможно доказать, поскольку отсутствуют сыворотки против указанного гипотетического аллеля К13.
Можно также предположить, что К13-эффект никак не связан с антигеном К13, а является проявлением пока неизвестного генетического процесса, приводящего к уменьшению синтеза антигенов Kell, в том числе антигена К13, который в этом случае не производится вовсе и его экспрессия соответствует 0.
Ожидаемые, но не встречающиеся гаплотипы KEL
Как известно, групповые антигены крови наследуются кодоминантно и независимо друг от друга. Однако существует много исключений, которые ставят под сомнение непреложность этого правила и еще раз подчеркивают огромное разнообразие не только форм групповых антигенов крови, но и способов их наследственной передачи. Приведем некоторые примеры:
- частичное доминирование (эпистазия) D над С в позиции цис. В серологических реакциях это проявляется следующим образом: эритроциты лиц CDe/cde (С и D цис) медленнее агглютинируются сывороткой анти-С и агглютинация выражена слабее, чем при тестировании эритроцитов лиц Cde/cDe (CnD транс);
- ослабление выраженности антигена D (Du) под действием гена С, расположенного по отношению к гену D в позиции транс. Этот феномен проявляется в том, что эритроциты лиц CDe/cde реагируют с сывороткой анти-D значительно сильнее, чем эритроциты лиц Cde/cDe;
- Кра-эффект - подавление синтеза антигенов Kell геном Кра; -К13-эффект - подавление синтеза антигенов Kell неизвестным геном, присутствующим в гаплотипе К13-.
Следует упомянуть еще об одном феномене - неравновесном сцеплении, которое проявляется в том, что антигены двух групповых систем предпочтительно наследуются вместе. Например, у людей, имеющих группу крови М, антиген S встречается в 2 раза чаще, чем у людей, имеющих группу крови N. Среди лиц М+ 72 % являются S+, среди N+ - 31 % S+. Считается, что ген S близко расположен к гену М, поэтому чаще наследуются вместе с ним, чем с геном N. Неравновесное сцепление генов Ми S по сравнению cNnSпослужило основанием полагать, что антигены MN и Ss представляют собой тесно связанную антигенную систему MNS.
В наследственной передаче антигенов Kell также проявляются свои особенности. Помимо Кра- и К13-эффекта существует еще одна необычная генетическая закономерность, а именно отсутствие Kell-гаплотипов, которые по логике вещей должны присутствовать. Речь идет о гаплотипах, кодирующих два редких антигена Kell и более, например: КраК, KJsa, KpaJsa, ККр° и др. Такие гаплотипы до сих пор не обнаружены.
Фенотипы, несущие два редких антигена, встречаются, например:
K+k+Kp(a+b+), K+k+Js(a+b+), K+k+Ul(a+)Kp(a-b+). Однако во всех случаях семейных исследований [154, 249, 251, 252, 288, 357, 402] установлено, что лица с фенотипом K+k+Kp(a+b+) генетически являются KKpb/kKpa, лица с фенотипом K+k+Js(a+b+) имеют генотип KJsb/kJsa, а лица с фенотипом K+k+Ul(a+)Kp(a-b+)- генотип KKpb/kUla.
Возникает вопрос: почему гены К и Кра, К и Jsa9 К и UIa, а также гены других редких Kell-антигенов не наследуются по два и более в одном гаплотипе?
До настоящего времени стройной генетической концепции, объясняющей это явление, не предложено. Некоторые авторы разделяют точку зрения Chown о том, что kKpbJsb представляет собой эволюционно более ранний гаплотип, который в результате единичных мутаций трансформировался в гаплотипы KKpbJsb, kKpaJsb, kKpcJsb и kKpbJsa. Далее логика нарушается, поскольку остальные 5 редких генов: Ula, Wka, К23, К24 и К25 - не имеют аллелей. Согласно предположению Chown, гаплотипы ККра, KJsa и KpaJsa могли сформироваться вследствие двойных мутаций, вероятность которых ничтожно мала в связи с редкостью генов К, Кра, Крс, Jsanjsp. Однако, как полагают Issitt и Anstee [204], с момента открытия редких антигенов Kell выполнено огромное количество исследований, и если бы такие двойные мутации (мутации предыдущих мутаций) существовали, они были бы обнаружены.
Не лишена оснований и другая точка зрения, что антигены системы Kell кодируются 10 гаплотипами (рис. 5.4). Один из них - общий (частый), 9 других -редкие. Первый гаплотип (общий) состоит из гена R- (R-минус), кодирующего все антигены Kell, за исключением редких. Другие 9 гаплотипов представлены одним из редких генов, которые кодируют 1 редкий и все остальные часто встречающиеся антигены.
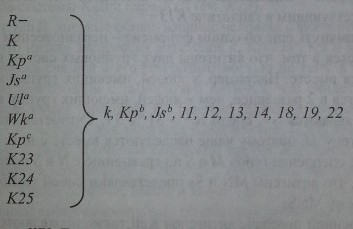
Гаплотипы KEL. Гаплотип 7?- производит общие Kell-антигены: k, Kpb, Jsb, 11, 12, 13,14,18,19 и 22; гаплотип К- антиген К и общие Kell-антигены; гаплотип Кра - антиген Краи общие Kell-антигены и т. д.
В соответствии с этой схемой гаплотипы, содержащие 2 редких антигена и более, исключены. Остается непонятным, почему в локусе KEL отсутствуют генные конверсии и другие формы обмена генетическим материалом между гомологичными хромосомами, как это имеет место в системе резус. По-видимому, в этом проявляется своеобразие рассматриваемого генного локуса.
Итак, подавляющее большинство людей гомозиготны по гаплотипу ШВ и соответственно содержат весь набор общих Kell-антигенов: k Kpb Jsb 11 12 13 14
|
Экзон |
Кодон |
Размер, к |
Кодируемые домены и антигены |
|
|
1 |
5'нто Metl |
0,34 |
|
|
|
2 |
2-27 |
0,29 |
Интрацеллюлярный домен |
|
|
а 5 |
28-74 |
0,26 |
Трансмембранный домен |
|
|
4 |
75-133 |
-2,6 |
К18 |
Экстрацеллюлярный домен |
|
5 |
134-175 |
0,33 |
|
|
|
6 |
176-224 |
-3,2 |
К14/К24/К/к |
|
|
7 |
225-245 |
0,093 |
|
|
|
8 |
246-308 |
0,23 |
Кра/Крь/Крс;К11/К17 |
|
|
9 |
309-358 |
-1,3 |
К22 |
|
|
10 |
359^101 |
-6 |
К23 |
|
|
11 |
402-438 |
-1,6 |
тои |
Экстрацеллюлярный домен |
|
12 |
439^171 |
0,24 |
|
|
|
13 |
472-497 |
0,44 |
К19,1Ла |
|
|
14 |
498-531 |
0,19 |
|
|
|
15 |
532-568 |
0,15 |
К12 |
|
|
16 |
569-590 |
0,23 |
HELLH |
|
|
17 |
591-647 |
0,35 |
Jsa/Jsb |
|
|
18 |
648-679 |
-1,3 |
|
|
|
19 |
680-732 3'нто |
|
|
|
Кра-эффект
lien и Lewis [87], открывшие антиген Кра и изучившие характер его наследования, обратили внимание на тот факт, что антиген к слабо выражен у некоторых членов семьи, имевших фенотип Кр(а+). На эритроцитах K-k+wKp(a+b~) антиген к был подавлен до такой степени, что его присутствие могло остаться незамеченным, если бы не были применены сильные анти-к-сыворотки. На эритроцитах K-k+Kp(a-b+) экспрессия к была в нормае.
Объяснение этому феномену нашли Race и Sanger [318], показав, что угнетающее действие гена Кра распространяется не только на к, но и на другие антигены системы Kell.
![Результаты исследования семьи, показывающие депрессивный эффект Кра на k (по Walsh и соавт. [390]). Ослабление к наблюдали у I м (Кра/К°) и П 2 (Кра/К).](/images/stories/stat/ad3.jpg)
Результаты исследования семьи, показывающие депрессивный эффект Кра на k (по Walsh и соавт. [390]). Ослабление к наблюдали у I м (Кра/К°) и П 2 (Кра/К).
В более поздних исследованиях было подтверждено супрессирующее влияние гена Кра на другие гены KEL, расположенные в позиции цис. Опубликованные данные свидетельствуют об уменьшенной экспрессии антигенов Ku (Ford [164]), Jsb (Ford [164], Tippett [370]), K18 (Barrasso и соавт. [98]), K19 (Sabo и соавт. [337]), TOU (Marsh и соавт. [272]) и других часто встречающихся антигенов Kell и napa-Kell (Walsh и соавт. [390]).
Manny и соавт. [256] сообщили, что экспрессия антигена К22 на эритроцитах Кр(а+Ь-) снижена по сравнению с эритроцитами Kp(a-b+). Ваг Shany и соавт. [96], напротив, констатировали, что экспрессия антигена К22 на эритроцитах Кр(а+Ь-) и Кр(а-Ь+) одинакова. Подавляющее действие гена Кра на другие гены KEL не всегда выявляют при обычном серологическом исследовании.
Впоследствии выяснилось, что супрессирующий Кра-эффект можно отчетливо наблюдать только при одном из трех обязательных условий:
- гомозиготность по гену Кра;
- присутствие гена К в позиции транс;
- присутствие молчащего гаплотипа К.
Кра-эффект сильнее проявляется при 3-м варианте, когда лица Кр(а+) имеют генотип Кра/К° [164, 390]. Присутствие гаплотипа К на противоположной хромосоме позволяет более четко увидеть подавляющее действие гена Кра на другие гены KEL в Х/?*-гаплотипе. Подобно этому подавляющее действие гена Кра на экспрессию антигена к и других Kell-антигенов легче обнаружить у людей kKpa/KKpby чем kKpa/kKp\ т. е. когда гены КраиК находятся в положении транс.
На рис. 5.3 представлены результаты семейного исследования, демонстрирующие супрессивный эффект гена Кра на ген к. Ослабление антигена k (к+w) наблюдалось у представителя первого (I м) и второго (II 2) поколения: I м имела генотип Кра/К°, II 2 - генотип Кра/К (Кра транс по отношению к К). У I м все высокочастотные антигены были супрессированы. У других членов семьи антигены Kell имели нормальную выраженность.
Tippett [370] установила, что сыворотки анти-k и aHra-Jsb имели разный титр при титровании 5 образцами эритроцитов Кр(а+Ь-). Один образец эритроцитов от человека k+ Kp(a+b~) Js(a-b+), второй гаплотип которого был, по-видимому, К, показывал отчетливо низкий титр; другие 4 образца k+ Kp(a+b-) Js(a4)+), полученные от гомозигот kKpaJsb/kKpaJsb, показали небольшое снижение титра антител анти-k и aHTH-Jsb, что свидетельствовало о сниженной экспрессии антигенов к и Jsb на эритроцитах гетерозигот К°.
Daniels и соавт. [150], Yazdanbakhsh и соавт. [404] полагают, что Кра-эффект проявляется только низкой синтезирующей способностью Хра-аллеля, в результате чего количество гликопротеинов Kell на эритроцитах снижено. Конформационные изменения в Kell-гликопротеине, которые могли бы препятствовать связыванию Kell-антигенов с антителами, не столь выражены.
Waheed, Kennedy [385] на модели экспрессии кДНК-конструктов в клетках эмбриональной почки человека нашли, что Кра (Тгр 281) замедляет продвижение Kell-гликопротеинов к поверхности клетки, удерживая их на уровне аппарата Гольджи. Для того чтобы обеспечить более эффективное продвижение Kell-гликопротеинов к клеточной поверхности, возникли мутации: часто встречающаяся - Kpb (Arg 281) и редко встречающаяся - Крс (Glu 281).
Daniels и соавт. [150] описали фенотип Ко, который на самом деле таковым не являлся. Несмотря на то что эритроциты этого человека не содержали Kell-антигенов, тем не менее они адсорбировали анти-к-антитела, которые затем обнаруживали в элюате. Кроме того, на эритроцитах пробанда отсутствовал антиген Кх, наличие которого является характерным признаком Ко-фенотипа. Наконец, 3 детей пробанда были Кр(а+Ь+), и маловероятно, что пробанд имел ген К°. Анализ экзона 8 показал, что пробанд был гомозиготен по гену Кр". Авторы отметили также мутации в Ж-гене, что позволило в итоге диагностировать у пробанда фенотип McLeod. Как считают авторы, Kell-ингибирующий эффект, приведший к Ко-подобному фенотипу, был обусловлен двумя причинами: гомозиготностью по гену Краи дефицитом Кх-протеина.
Необходимо отметить, что ^"-эффект проявляется только в отношении часто встречающихся Kell-антигенов. Редкие антигены Kell (имеющие частоту менее 10 %): К, Jsa, Кра, Крс и др. - не подвержены ингибиции, поскольку ген Краи гены К, Jsa, Кра, Крс в одном гаплотипе, в положении цис, не встречаются (см. Ожидаемые, но не встречающиеся гаплотипы KEL).
Kikuchi и соавт. [225] привели данные, показывающие, что в отличие от гена Кра его редко встречающийся аллель Крс не влияет на гены KEL, с которыми он находится в положении цис, как это имеет место при Кра-эффекте. Авторы не отметили подавления экспрессии антигенов k, Jsb и других часто встречающихся антигенов Kell и napa-Kell на эритроцитах лиц с фенотипом Кр(а-Ь-с+).
К выводу о том, что локус KEL расположен на хромосоме 7, пришли Zelinski и соавт. [406], Parsons и соавт. [303]. Авторы обнаружили связь между генами KEL и геном пролактининдуцирующего протеина (PIP), который локализован на хромосоме 7 в позиции q32-q36.
Purohit и соавт. [315] нашли, что локус KEL связан с геном цистофиброза, который, так же как PIP, локализован на длинном плече хромосомы 7.
Последующие исследования, проведенные Murphy и соавт. [295], Lee и соавт. [244] с помощью методов гибридизации in situ и метода клонирования Kell-специфической кДНК, подтвердили расположение генов KEL на участке 7q33-7q35.
Генный комплекс Kell включает 19 экзонов и занимает примерно 21,5 кб (Lee и соавт. [242]).
Экзон 1 содержит 5!-нетранслируемую область, кодон метионина 1, инициирующий трансляцию, SP1- и GATA-1 -связывающие участки. Как полагают Camara-Clayette и соавт. [119], экзон 1 вовлечен в отрицательную регуляцию промотора в неэритроидных тканях.
Экзон 2 кодирует цитоплазматический домен и, возможно, второй инициирующий трансляцию метионин 20.
Экзон 3 кодирует трансмембранную, наиболее короткую часть Kell-пожпептида, которая состоит примерно из 20 аминокислотных остатков (см. рис. 5.2).
Экзоны 4-19 ответственны за экстрацеллюлярный, самый протяженный домен. Аминокислотная последовательность, придающая Kell-протеину свойства цинкзависимой металлопротеиназы, кодируется экзоном 16, который, по данным Lee и соавт. [242], на 54,5 % идентичен эквивалентному участку гена NEP, кодирующего вазоконстриктор неприлизин.
Lee и соавт. [242] нашли, что 5'-латеральная область от -176 до -1 нуклеоти-да содержит три участка, связывающих GATA-1. По мнению Shivdasani и Orkin
м. ZU4], GATA-1 является транскрипционным фактором, необходимым для формирования эритроидных тканей.
Мутации в участке связывания GATA-1 приводят к нарушению синтеза на эритроцитах гликопротеина Duffy; эти мутации наблюдают у большинства людей с фенотипом Fy(a-b-). Неизвестно, определяют ли эти мутации отсутствие белка Kell на эритроцитах людей с фенотипом Ко.
Lee и соавт. [242] полагают, что локус KEL регулируется эритроидными факторами транскрипции.
Организация гена KEL*
|
Экзон |
Кодон |
Размер, кб |
Кодируемые домены и антигены |
|
|
1 |
5'нто Metl |
0,34 |
|
|
|
2 |
2-27 |
0,29 |
Интрацеллюлярный домен |
|
|
а 5 |
28-74 |
0,26 |
Трансмембранный домен |
|
|
4 |
75-133 |
-2,6 |
К18 |
Экстрацеллюлярный домен |
|
5 |
134-175 |
0,33 |
|
|
|
6 |
176-224 |
-3,2 |
К14/К24/К/к |
|
|
7 |
225-245 |
0,093 |
|
|
|
8 |
246-308 |
0,23 |
Кра/Крь/Крс;К11/К17 |
|
|
9 |
309-358 |
-1,3 |
К22 |
|
|
10 |
359^101 |
-6 |
К23 |
|
|
11 |
402-438 |
-1,6 |
тои |
Экстрацеллюлярный домен |
|
12 |
439^171 |
0,24 |
|
|
|
13 |
472-497 |
0,44 |
К19,1Ла |
|
|
14 |
498-531 |
0,19 |
|
|
|
15 |
532-568 |
0,15 |
К12 |
|
|
16 |
569-590 |
0,23 |
HELLH |
|
|
17 |
591-647 |
0,35 |
Jsa/Jsb |
|
|
18 |
648-679 |
-1,3 |
|
|
|
19 |
680-732 3'нто |
|
|
|
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}