



С 1951 г. по 1963 г. опубликована серия исследований, показавших, что лица D+, так же как D-, могут вырабатывать анти-О-антитела [216, 386, 446, 614, 645, 658, 671], реагирующие со всеми образцами эритроцитов D+, за исключением собственных. Авторы подчеркивали, что антиген D мозаичен и представлен на эритроцитах многими эпитопами, которые могут выступать в качестве самостоятельных иммуногенов.
Обычно эпитопы присутствуют на эритроцитах людей D+ в полном составе. Люди, имеющие неполный комплект эпитопов, способны вырабатывать антитела к отсутствующим у них компонентам. Фенотип таких людей назван частичным, или парциальным, D.
При адсорбции сывороток анти-D, полученных от людей D-, эритроцитами разных категорий частичного D не удавалось выделить фракции, способные распознавать отдельные эпитопы D-антигена. Элюаты, как правило, содержали антитела, идентичные тем, что находились в сыворотке изначально. Другими словами, антитела анти-D не сепарировались на отдельные эпитопные специфичности и представляли собой единую рецепторную структуру, направленную одновременно против множества D-эпитопов.
Антитела анти-D, присутствующие у лиц с парциальным D, имеют более узкий диапазон реагирования, чем анти-О-антитела лиц D-, поскольку в них отсутствуют рецепторы к отдельным эпитопам D. Иными словами, антитела, продуцируемые людьми с парциальным D, также являются парциальными.
По характеру перекрестного реагирования парциальные антитела разделили на группы.
Wiener, Unger и другие исследователи [216, 614, 645, 671, 716 ], установившие, что Rho-aHTHreH состоит из 4 компонентов (RhA, RhB, Rhc и RhD), предложили классификацию RhQalPhabet. Если один из компонентов в фенотипе присутствовал, его обозначали индексом с заглавной буквой, если отсутствовал -строчной буквой. Таким образом, эритроциты фенотипа Rh^^ содержали компоненты RhA и RhB, а компоненты Rhc и RhD в них отсутствовали. Как обнаружили исследователи, антитела, полученные от человека с эритроцитами RhA, реагировали с нормальными эритроцитами Rho и эритроцитами Rha, но не реагировали с эритроцитами RhA. Антитела, продуцируемые человеком, имевшим эритроциты RhB, реагировали с эритроцитами Rho и эритроцитами Rhb, но не реагировали с эритроцитами RhB и т. д.
В настоящее время алфавитную классификацию Rhoalphabet не используют и она упоминается лишь как имеющая научно-познавательное значение.
Классификация по категориям, существенно расширенная в настоящее время, предложена Tippett и Sanger [657, 661].
Первая ее версия опубликована в 1962 и 1963 гг. Суммируя результаты исследования сначала 18, а затем 29 сывороток анти-D, полученных от лиц с парциальными D-антигенами, авторы выделили 6 категорий антигенов, отличающихся неодинаковым перекрестным реагированием (табл. 4.12).
КI категории D-антигенов был отнесен обычный D-антиген. Люди с антигеном категории DI анти-О-антител не вырабатывают.
Ко II категории отнесен антиген D, содержащийся у людей, которые продуцируют антитела против D-антигенов I, III, IV и V категории. К D-антигенам II и VI категории такие люди антител не вырабатывают.
К III категории отнесен антиген D, встречающийся у лиц, продуцирующих антитела против D-антигенов I, II и IV категории.
Антиген D IV категории отличается от антигена D лиц первых трех категорий. Люди, имеющие этот антиген, способны вырабатывать антитела к D-антигенам всех категорий, за исключением IV.
Отличительные признаки D-антигенов V и VI категории сводятся к тому, что люди, имеющие эти антигены, способны продуцировать антитела к антигенам D других категорий, но лишены этой способности в отношении собственного D-антигена.
Эритроциты, содержащие антигены D разных категорий, различаются по частоте реагирования со стандартными сыворотками анти-D, полученными от людей D-. Образцы эритроцитов, отнесенные к категории IV, агглютинировались 96 % сывороток анти-D, категории V - 74 %. Эритроциты категории VI реагировали только с 35 % сывороток анти-D.
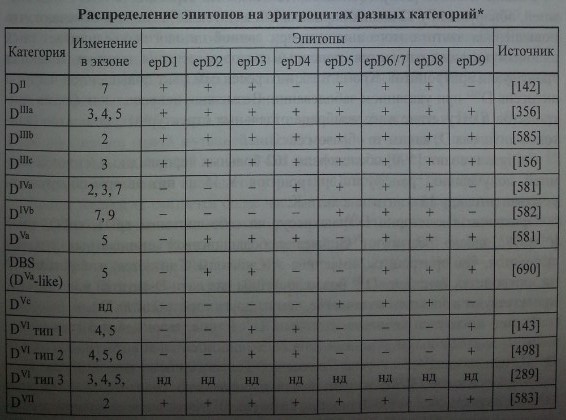
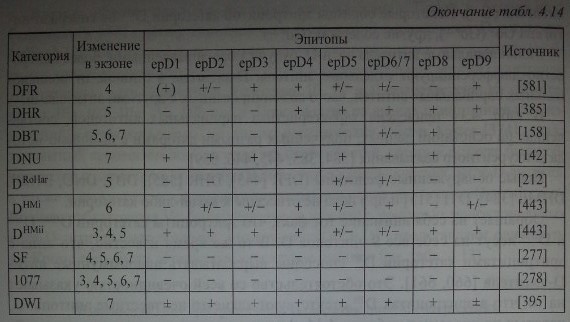
Из данных, представленных в табл. 4.12, можно заключить, что не все эпитопы D одинаково иммуногенны. По-видимому, эпитопы, отсутствующие на эритроцитах DVI, как раз и являются более иммуногенными, поскольку их отсутствие обусловливает низкий процент реагирующих с этими эритроцитами сывороток. Напротив, высокая частота реагирования Dn, DIH, DIVсвидетельствует о том, что они лишены менее иммуногенных эпитопов. Если судить по данным табл. 4.14, к более иммуногенным следует отнести все эпитопы от epDl до epD9, а к менее иммуногенным - те, которые «прячутся» за знаком минус (отсутствие сильного эпитопа).
Вторую, обновленную версию своей классификации Tippett и Sanger опубликовали в 1977 г. [660]. Она включала 5 категорий парциальных D-антигенов (табл. 4.13). Категория I была исключена, так как выяснилось, что некоторые образцы крови, отнесенные к этой категории в первоначальной версии, содержали транзиторные анти-О-антитела. Категории III, IV и V подразделены на субкатегории: Ша, Illb, Шс; IVa, IVb и т. д. Новая версия не внесла принципиальных изменений в первоначальную, но, как комментировали Issitt и Anstee [374]: «дала элегантный серологический прогноз тех особенностей, которые позднее были обнаружены с помощью моноклональных эпитопспецифических aHTH-D-антител, а также в результате исследования генов, кодирующих частичные D-антигены». Позднее субкатегория Vc была упразднена, так как с помощью моноклональных aHTH-D-антител было показано, что антигены DVc и Divb представляют одну группу. Кроме того, были детализированы другие позиции (см. табл. 4.13).
Таблица 4.13
Взаимодействие эритроцитов и сывороток людей, содержащих парциальные D-антигены и парциальные анти-О-антитела*
|
Категория эритроцитов |
Реакция анти-В-антител, полученных от лиц, имеющих эритроциты категории |
|||||||||||
|
II |
Ша |
ШЬ |
Шс |
IVal |
IVa2 |
IVb |
Va |
Vb |
Vc |
Vli |
Vlii |
|
|
II |
— |
+ |
+ |
+ |
+ |
+ |
± |
+ |
+ |
I |
+ |
+ |
|
Ша |
|
— |
+ |
— |
|
+ |
+ |
|
+ |
+ |
|
+ |
|
ШЬ |
+ |
— |
— |
— |
+ |
+ |
+ |
± |
|
+ |
+ |
+ |
|
Шс |
|
— |
4- |
— |
+ |
+ |
+ |
+ |
+ |
+ |
|
+ |
|
IVa |
— |
1 |
|
— |
— |
— |
— |
+ |
+ |
— |
+ |
+ |
|
IVb |
— |
1 |
|
|
— |
— |
— |
— |
— |
1 |
+ |
— |
|
Va |
+ |
— |
|
— |
+ |
|
+ |
— |
± |
— |
+ |
— |
|
Vb |
+ |
— |
|
— |
+ |
— |
— |
— |
— |
— |
— |
—■ |
|
Vc |
+ |
— |
+ |
— |
| |
— |
|
— |
— |
— |
|
— |
|
VI |
— |
— |
+ |
— |
+ |
— |
+ |
— |
— |
— |
— |
— |
* По Tippett, Sanger [660] и другим источникам.
Лица, имеющие эритроциты категории DHI (a, b и с), содержат 2 разных анти-О-антитела. Об этом свидетельствует тот факт, что эритроциты D11, D111 и других категорий реагируют с этими сыворотками неодинаково. В частности, анти-О-антитела людей DIIIb агглютинируют эритроциты D11, D1113"^. В то же время эритроциты DIIIa и Dlllc сыворотками лиц DIIIb практически не агглютинируются. Такая же двойная специфичность характерна для антител, продуцируемых лицами DVI. Так, лица и D™' продуцируют 2 типа антител, один из которых реагирует с эритроцитами DIVb, DVa и DVc, а другой с этими эритроцитами не реагирует. Анти-О-антитела, вырабатываемые лицами DIVa и DIvb практически однотипны, так же как и антитела, вырабатываемые лицами DVa, Dvb и DVc.
Эритроциты DIIIb представлены двумя группами. К одной группе относят эритроциты, содержащие антиген G (G+), к другой Ё не содержащие этого антигена (G-). Некоторые образцы эритроцитов категории DIVa не содержат антигена Goa (Go8-), другие содержат (Goa+). Щ'
Моноклональные антитела анти-D, будучи узконаправленными, более контро-астно дифференцируют отдельные эпитопы D-антигена в отличие от поликло-нальных парциальных антител, часто комбинированных с другими антителами.
В табл. 4.14 приведена характеристика парциальных антигенов, составленная на основании данных, полученных разными авторами с помощью метода конкурентного связывания [384,385,445,446,659].
Новые обнаруженные фенотипы (DFR [445], DHR [385], DBT, DNU, DHMi, DHMii [384], DTI [513] и др.) пока не отнесены к какой-либо категории.
Обращает на себя внимание тот факт, что эритроциты категории DIIIa, DIIIbи DIIIC несут на себе все 9 эпитопов D. В то же время известно, что лица с эритроцитами категории D111 способны вырабатывать аллоиммунные анти-О-антитела [660, 661]. Это обстоятельство со всей очевидностью указывает на то, что в эритроцитах D111 отсутствуют какие-то неизвестные эпитопы D, которые не внесены в таблицу 4.14. Антитела к этим эпитопам пока не найдены. Аналогично ведут себя эритроциты категории Dn,DIV, DvhDVI1, которые в одних случаях реагируют с сыворотками анти-D, а в других не реагируют в зависимости от совпадения специфичности парциальных антигенов и парциальных антител.
Castilho и соавт. [198], обследовав 130 больных серповидно-клеточной анемией, обнаружили у 12 из них парциальный антиген DIIIa. Авторы отмечают, что повышенная частота этого антигена при данной патологии увеличивает риск аллоиммунизации пациентов, основным методом лечения которых являются трансфузии эритроцитов. Кроме того, интересен сам факт повышенной частоты антигена DIIIa при указанном заболевании. Возможно, неправильная форма эритроцитов и отсутствие в их мембране отдельных структурных элементов (эпитопов антигена D) каким-то образом связаны.
Calligas и соавт. [190] обследовали 162 больных серповидно-клеточной анемией, получавших трансфузии эритроцитов. У 15 из них авторы обнаружили резус-антитела, у 13 - антитела анти-К.
Эритроциты категории DIVa не несут на себе эпитопов epDl, epD2, epD3 и epD9, однако реагируют сильнее с большинством стандартных анти-D-сывороток, чем эритроциты, имеющие эти эпитопы. Считается, что эпитопы D на эритроцитах категории DIVa более доступны для анти-Б-антител и, возможно, присутствуют в большем количестве, чем на других эритроцитах D+.
У лиц, носителей антигена категории DVI I типа, экзоны 4 и 5 гена сЕ замещены аналогичными экзонами гена D. У лиц, носителей антигена категории DVIII типа, экзоны 4, 5 и 6 гена Се замещены аналогичными экзонами гена D.
Wagner и соавт. [690] нашли новый парциальный D-антиген - DBS, или Dv-like, являющийся разновидностью антигена DVa. Этот антиген |содируется ги-^щ^фт геном RHD-R^^(5)-RHD с делецией интрона 4 и частщэкзона 5. На эритроцитах Dv-like отсутствуют эпитопы epD4, ерШ2, epD17, epD18 и epD22, которые обычно присутствуют на эритроцитах DVa. Указанное отличие антигена Dv-like от антигена DVa было ассоциировано только с одной нуклеотидной заменой А 226 Р, которая, видимо, и привела к выпадению некоторых эпитопов. Ранее Wagner и соавт. [691] описали разновидности парциальных антигенов Divb-like, DNU-like и DFR-like, различающиеся точками мутаций в RHD-тене и отсутствием некоторых эпитопов (табл. 4.15). По данным Zhou и соавт. [729], полиморфизм антигена DVa обусловлен гибридным геном RHD/CE, в котором экзон 5 и интрон 5 гена D замещен гомологичными последовательностями гена СЕ.
Таблица 4.15
Разновидности парциальных антигенов DIV, DNB, DFR
|
Фенотип (категория парциального D-антигена) |
Количеств о случаев |
Нуклеотидная замена в гене RHD |
Недостающие эпитопы D на эритроцитах |
|
D^-like (D™ IV типа) |
3 |
D350 m Н, G353 ^W,A354-+N |
1-6,23,31 34-36 |
|
DNU-like (DNB) |
2 |
G355 Ш S |
6,31 |
|
DFR-like (DFR) |
1 |
Н166 Р |
10,11,22,31,32 |
Flegel и соавт. [288,289] обнаружили III тип парциального антигена D^. В категории III типа экзоны 3, 4, 5 и 6 гена D замещены эквивалентными частями гена Се. Авторы нашли, что число антигенных участков на 1 клетку для разных типов антигена составляет: для I типа - 500, для II типа - 2400 и для
Ш типа - 12 000. Таким образом, эритроциты, несущие антиген III типа, содержат нормальное для эритроцитов D+ количество антигенных участков.
Два типа эритроцитов (I и III) содержат антиген BARC (Rh52), эритроциты II типа не содержат этого антигена.
Jones и соавт. [385] описали новый парциальный антиген - DHR. Фенотип DHR обусловлен одной мутацией - G 686 —► А - в экзоне 5 гена Д которая приводит к замене Arg 229 —► Lis в 4-й экзофациальной петле полипептида D. Аминокислотная замена приводит к утрате epD 1, epD2, epD 12 и epD20.
Генная конверсия считается более частым механизмом появления парциального D-фенотипа. Однако есть несколько примеров простых замен, обусловливающих парциальные антигены D различных категорий: Dm - Leu ПО —► Pro [589, 694], D"-Ala 354 У Asp [144], DNU - Gly 353 Л Arg [144], DHMi - Thr 285 -* He [384].
В 2004 г. Kormoczi и соавт. [395] описали парциальный D-антиген, названный ими DWI. Он был обнаружен у 74-летней жительницы Восточной Австрии, имевшей группу крови 0(1) Rh+. Сыворотка женщины содержала анти-О-антитела, что послужило основанием заподозрить у нее парциальный вариант D. В анамнезе имелись 2 беременности от мужа D+ и несколько ге-мотрансфузий в связи с гинекологической операцией. Женщина и 2 ее родственницы (сестра и племянница) имели генотип DWICe/cde. На эритроцитах
DWI+ отсутствовали редкие антигены ЦЦ V9 VS, Dw, Goa, Evans, Tar, Riv, FPTT, BARC, JAL, которые ассоциированы с парциальными антигенами D, Сс или Ее. Антигенная емкость эритроцитов DWI+ составила 8000-8600 антигенных участков на 1 эритроцит. При семейном исследовании установлено, что антиген DWI передается с гаплотипом DWICe, но не с DWIce. При эпитоп-ном картировании зафиксирована незначительная модификация антигена D, проявляющаяся в виде ослабленной экспрессии эпитопов Dl.l, D9.1 и D16.1. При ДНК-типировании авторы нашли в экзоне 7 гена RHD простую нуклео-тидную замену Т 1073 —> С, приводившую к замещению Met 358 —> Thr в 6-й экстрацеллюлярной петле D-полипептида.
Эритроциты DWI+ не агглютинировались одной из 79 моноклональных анти-D-cbmopoTOK, хорошо реагировавших с другими фенотипами D+. Авторы применили эту сыворотку для отсева лиц D+, с тем чтобы среди оставшихся нереагирующих или слабореагирующих образцов найти DWI+. Однако среди 2288 резус-положительных австрийцев антиген DWI выявлен не был, за исключением трех упомянутых выше женщин.
Вначале была предложена 9-эпитопная модель парциальных D-антигенов. Специально отобранные моноклональные антитела дифференцировали 8 эпитопов антигена D (epDl-epD9). Эпитопы D6 и D7 были объединены в epD6/7, так как серологические реакции эритроцитов, несущих эти эпитопы, оказались практически одинаковыми.
Faas и соавт. [276] предложили 15-эпитопную модель, в которой эпитопы epDl, epD2, epD5, epD6/7 и epD9 подразделяют на субэпитопы (Dl.l, D1.2; D5.1, D5.2; D9.1, D9.2 и т. д.) - от 2 до 10 субэпитопов в каждом эпитопе. Кроме того, авторами установлено 6 новых эпитопов, получивших номера с 10 по 15.
Jones и соавт. [386] разработали модель, включающую 30 серологически различимых эпитопов D (табл. 4.16). Как видим, эпитопная конструкция антигена D, сложившаяся к настоящему времени, еще далека от своего завершения.


{kind=link}