Поиск по сайту
- Вы здесь:
-
Главная

-
book
-
Группы крови человека
- Система Duffy
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...

- Категория: Система Duffy
Малярийный паразит Plasmodium vivax вызывает четырехдневную форму малярии, которая протекает более легко, чем заболевание, вызываемое Plasmodium falciparum. Большинство негроидов не подвержены малярии, вызываемой Р vivax.
Miller и соавт. [109, ПО], а затем Hadley и соавт. [62, 63] установили, что эритроциты Fy(a-b-) in vitro невосприимчивы к инвазии обезьяньими малярийны-ми паразитами Plasmodium knowlesi. До Miller и соавт. инвазию эритроцитов не связывали с фенотипом клеток по системе Duffy. Это стало очевидным после проведения аналогичных экспериментов с Р vivax (Barnwell и соавт. [17]).
Среди искусственно зараженных P. vivax 17 добровольцев (11 негров и 6 белых) симптомы заболевания развились у всех, за исключением 5 негров с фенотипом Fy(a-b-) (Miller и соавт. [109]).
Среди 420 жителей Гондураса, из которых 247 имели фенотип Fy(a-b-), 14 (7 негров и 7 белых) болели малярией, вызванной P. vivax. Все 14 были Duffy-положительными (Spencer и соавт. [152]).
Антитела к P. vivax обнаруживали исключительно у Duffy-положительных лиц, а антитела к P. falciparum - с одинаковой частотой у Duffy-положительных и Duffy-отрицательных лиц (Spencer и соавт. [152]).
Весьма вероятно, что высокая частота фенотипа Fy(a-b-) среди жителей Африки является результатом естественного отбора особей, устойчивых к инфицированию Р vivax. Лица, гомозиготные по молчащему гену Fy, имеют селективное преимущество, поскольку не болеют малярией. В некоторых районах Западной Африки частота гена Fy среди населения достигает почти 100 %, хотя Р vivax в этих областях не встречается (Welch и соавт. [176]). Элиминацию паразитов в этих районах объясняют отсутствием подходящего объекта (людей Fy+) для реализации жизненного цикла плазмодия.
Эритроциты Fy(a-b-) устойчивы in vitro к инвазии P. vivax и Р knowlesi, в то время как в эритроциты Fy(a+b+) указанные малярийные плазмодии проникают (Barnwell и соавт. [17], Miller и соавт. [108, ПО]). Устойчивость эритроцитов Fy(a-b-) представителей других рас (индейцы, австралийцы) к инвазии P. knowlesi не изучена.
Малярийные плазмодии P. knowlesi способны связаться с эритроцитами Fy(a-b-), однако не могут проникать внутрь (Mason и соавт. [102], Miller и соавт. [107, ПО]). Инвазия Duffy-положительных эритроцитов P. vivax и Р knowlesi блокируется моноклональными антителами анти-Руб (Barnwell и соавт. [17], Miller и соавт [ПО]).
Хемокины IL-8 и MGSA, для которых Duffy-гликопротеин является рецептором, также блокировали инвазию Duffy-положительных эритроцитов P. knowlesi [10]. Обработка указанных клеток химотрипсином повышала их устойчивость к инвазии Р vivax и Р knowlesi, а трипсин подобного эффекта не оказывал (Barnwell [17], Miller и соавт. [ПО]). Эффект оказался сходным в действии протеаз на антигены Fya, Fyb и I но не Fy3 и Fy5, которые были устойчивы к действию химотрипсина. Интересно, что эритроциты Fy(a-b-), обработанные
трипсином или сиалидазой, поддавались инвазии P. knowlesi (Mason и соавт. [102]). Указанные малярийные паразиты внедрялись преимущественно в рети-кулоциты, несущие повышенное количество антигена Fy6 по сравнению со зрелыми эритроцитами (Woolley и соавт. [181]).
В табл. 10.6 приведены Duffy-фенотипы низших приматов, эритроциты которых подвержены инвазии P. vivax и P. knowlesi. Несмотря на отсутствие антигена Fy6 (антигены Fyb и Fy3 присутствуют), эритроциты Macacus rhesus заражаются P. knowlesi, но резистентны к P. vivax. Эритроциты обезьян-капуцинов Нового Света имели фенотип Fy:-l,-2, 3, 6 и не поражались P. vivax и P. knowlesi. Эти данные свидетельствуют о важной роли эпитопов Fy6 в инвазии эритроцитов паразитами P. vivax (Palatnik, Rowe [130]).
Таблица 10.6.
Инвазия эритроцитов человека и обезьян малярийными плазмодиями
|
Эритроциты |
Реакция с сывороткой анти- |
Подверженность инвазии |
Связывание cDuffy-гликопротеинами |
||||||
|
Fy8 |
Fyb |
Fy3 |
Fy6 |
P knowlesi |
P vivax |
P. knowlesi |
R 1J vivax |
||
|
Человека |
Fy:l,2,2,6 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Fy:l,-2,2,6 |
|
— |
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Fy:-1,2,2,6 |
— |
|
+ |
+ |
+ |
+ |
+ |
+ |
|
|
Fy:-l,-2,-2,-6 |
— |
— |
— |
— |
— |
— |
— |
— |
|
|
Обработанные химотрипсином |
— |
— |
+ |
— |
— |
— |
— |
— |
|
|
Обезьян |
Масаса mulatta |
— |
+ |
+ |
— |
+ |
— |
+ |
— |
|
Saimiri sciureus |
— |
— |
+ |
+ |
+ |
+ |
+ |
— |
|
|
Cebus apelh |
— |
— |
+ |
— |
— |
— |
+ |
— |
|
|
Aotus triviratus |
— |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Протеины, связывающиеся с Duffy-положительными эритроцитами, но инертные по отношению к эритроцитам Fy(a-b-), обнаружены Haynes и соавт. [68], Wernheimer, Barnwell [177] в супернатантах культур Р vivax и Р knowlesi на этапе образования мерозоитов. Протеины, выделенные из Р knowlesi, и Р vivax, имели мол. массу 135 и 140 кДа соответственно. Связывание их с эритроцитами удавалось ингибировать антителами анти-Руб, а также хемокинами IL-8 и MGSA (Haynes и соавт. [68], Miller и соавт. [Ill], Wernheimer, Barnwell [177]). Обработка Duffy-положительных клеток химотрипсином предотвращала связывание |§рих протеинов с клетками, а обработка трипсином была эффективной. Эритроциты Fy(a-b-), подвергнутые инвазии P. knowlesi, не связывали протеины, выделенные из культур указанного паразита (Mason и соавт. [102], Haynes и соавт. [68]). Очищенный протеин паразита специфически связывался с очищенным Duffy-протеином. Гены, кодирующие синтез обоих протеинов, удалось клонировать. Установлено, что экстрацеллюлярные домены каждого из белков в шести регионах имели одинаковую аминокислотную последовательность (Adams и соавт. [9]). Клетки линии COS-7 экспрессировали участки, богатые цистеином, за счет которых формировались розетки с Duffy-положительными эритроцитами, но не эритроцитами Fy(a-b-) (Chitnis, Miller [33]). Розеткообразование блокировалось синтетическими пептидами, в кото-рьк аминокислотные последовательности в позициях 8-42 были идентичны таковым на N-терминальном домене Duffy-гликопротеина (Chitnis и соавт. [32]). Эритроциты лиц, гетерозиготных по мочащему гену Fy, связывали меньшее количество протеина Р. vivax по сравнению с клетками людей, обладающих двумя генами FY. Это может свидетельствовать о том, что в районах, эндемичных по P. vivax, даже гетерозиготность по гену Fy создает селективные преимущества (Michon и соавт. [106]). Протеины P. knowlesi и P. vivax, вступающие в реакцию соединения с Duffy-гликопротеином эритроцитов человека, имели высокую степень гомологии с гликопротеином P. falciparum (Adams и соавт. [9]). Анализ ли-гандов, связывающихся с Duffy-гликопротеинами, позволил идентифицировать вариабельные гены var, кодирующие эндотелиальные цитоадгезивные протеины у P. vivax (Hadley, Peiper [63], Pogo, Chaudhury [135]).
Singh и соавт. [149] выделили функциональный участок протеина P. vivax -PvRII, связывающегося с Duffy-гликопротеином. Протеин PvRII оказался высоко-иммуногенным и стимулировал выработку специфических антител в высоком титре. Последние ингибировали связывание P. vivax с эритроцитами. Протеин PvRII перспективен для создания вакцины против P. vivax, которая предохраняет также от заражения P. falciparum (Singh и соавт. [149], Williams и соавт. [180]).
Jilma-Stohlawetz и соавт. [82] сообщили о различиях в содержании хемокинов у мужчин и женщин. Содержание в плазме фактора хемотаксиса моноцитов было выше у мужчин, а также у лиц с фенотипом Fy(a-b-).
У реципиентов с пересаженной почкой во время кризов отторжения (Segerer и соавт. [145, 146], Alkalin, Neylan [12]), а также у больных раком предстательной железы (Lentsch [88]) концентрация хемокинов повышалась.
Н.Д. Герасимова [1] нашла повышенную частоту фенотипа Fy(a-) у онкологических больных.
- Категория: Система Duffy
Гликопротеины Duffy способны связываться с хемокинами, относящимися к факторам воспаления и хемотаксиса, поэтому их нередко обозначают как DARC (Duffy antigen receptor chemokine). Хемокины участвуют во многих межклеточных взаимодействиях, в том числе в активации лейкоцитов (Rollins [141]). Существует 3 больших класса хемокинов: С-Х-С, С-С и С, обозначенных так в связи с позицией цистеинового остатка в N-терминальном участке пептида. Большинство хемокино-вых рецепторов эритроцитов представлено семейством интегральных гликопротеинов. Трансмембранные гликопротеины G-типа представляют группу протеинсвязы-вающих рецепторов (Ji и соавт. [81], Murdoch, Finn [120]), воспринимающих экстра-целлюлярные сигналы через феромоны, нейромедиаторы и гормоны. Хемокиновые рецепторы специфичны к одному или нескольким гликопептидам.
Duffy-гликопротеины связываются с хемокинами С-Х-С и С-С (Darbonne и соавт. [45], Horuk и соавт. [71], Neote и соавт. [122]). Некоторые хемокины С-Х-С идентифицированы как интерлейкины-8 и факторы стимуляции роста меланомы (MGSA). Хемокины С-С участвуют в формировании рецепторов Т-клеток и продукции факторов хемотаксиса для моноцитов (RANTES, МСР-1). Гликопротеин Duffy не содержит рецепторов для хемокина С - фактора хемотаксиса лимфоцитов (Szabo и соавт. [153]). В отличие от других протеинов G-типа, в структуре Duffy-гликопротеина нет мотива Asp-Arg-Tyr (DRY) во втором цитоплазматическом домене, последний взаимодействует с протеином, связывающим гуанозинтрифосфат (Hadley, Peiper [63]).
Эритроциты лиц Fy(a-b-) не обладают способностью связывать хемокины (Horuk и соавт. [70], Tournamille и соавт. [162]), а эритроциты Fy(a-b+w) связывают некоторое количество указанных субстанций (Tournamille и соавт. [162], Zimmerman и соавт. [184]).
Хемокиновые рецепторы располагаются на второй и четвертой экстрацел-люлярной петле Duffy-гликопротеина вблизи одной из дисульфидных связей (Tournamille и соавт. [161, 163]).
Антитела к антигенам Fya, Fyb и Fy6, связывающиеся с соответствующими эпитопами Duffy-гликопротеина, и моноклональные антитела анти-РуЗ способны блокировать хемокины (Chaundhury и соавт. [31], Hausman и соавт. [67], Horuk и 70], Lu и соавт. [96], Szabo и соавт. [153], Tournamille и соавт. [163]). Интерлейкин-8 связывается с эритроцитами, обработанными трипсином, сиалидазой, N-гликаназой, но не связывается с эритроцитами, обработанными папаином и а-химотрипсином (Wasniowska и соавт. [171]). Два последних фермента расщепляют N-терминальный домен Duffy-гликопротеина.
Физиологические функции Duffy-гликопротеинов до конца не ясны. Высказывались предположения, что Duffy-гликопротеины эритроцитов адсорбируют избыточное количество воспалительных хемокинов (Darbonne и соавт. [45]) и интерлейкина-8, содержание которого повышается в плазме крови при инфаркте миокарда (de Winter и соавт. [47]). Однако в действительности эта функция эритроцитов вряд ли имеет столь существенное значение, поскольку у многих людей, особенно у негроидов, Duffy-гликопротеины на эритроцитах отсутствуют.
Как уже отмечалось выше, Duffy-гликопротеины содержатся и в неэритроидных клетках, в том числе у лиц Fy(a-b-), включая негроидов. Почечная изо-форма Duffy-гликопротеина связывала хемокины в той же степени, что и эри-троцитарная (Hadley и соавт. [61]).
Hadley и Peiper [63] полагают, что высокая консервативность гена FY, проявляющаяся в разных тканях, свидетельствует о важной роли Duffy-гликопротеинов в физиологии человека.
Клетки эритроидной линии К562, подвергнутые трансфекции кодирующей ДНК гена FY, связывали хемокины (Peiper и соавт. [132]). Duffy-гликопротеин присутствовал как трансмембранный структурный элемент в эндотелиальных клетках (Chaudhuri и соавт. [27]). Он может принимать участие в эндоцитозе, и возможно при этом инициирует синтез факторов хемотаксиса, вызывающих миграцию лейкоцитов (Hadley, Peiper [63], Lee и соавт. [87]). Существенное повышение содержания Duffy-гликопротеина отмечено в тканях почек ВИЧ-инфицированных лиц, а также больных уремией с гемолитическим синдромом. Последнее дает основание полагать, что Duffy-гликопротеины могут играть определенную роль в патогенезе воспалительных процессов в почечной ткани (Lu и соавт. [95]).
Гены, гомологичные генам FY человека, обнаружены у обезьян, коров, свиней, кроликов и мышей (Chaudhuri и соавт. [29], Hadley, Peiper [63], Luo и соавт. [97]). Эритроциты мышей связывали хемокины мыши и человека (Szymanski и соавт. [154]) так же, как и клетки эритроидной линии К562, подвергнутые трансфекции кДНК Dfy - мышиным гомологом гена FY человека (Luo и соавт. [97]). Мыши, дефицитные по гену Dfy, были соматически здоровы и их реакция на инъекцию воспалительных хемокинов не отличалась от таковой у обычных животных (Dawson и соавт. [46], Luo и соавт. [98]).
Некоторые лица Fy(a-b-), гомозиготные по мутации, инактивирующей ген FY, соматически здоровы, несмотря на полное отсутствие в их тканях Duffy-гликопротеинов. Вероятно, при отсутствии Duffy-гликопротеинов их биологическую функцию могут выполнять другие структуры (Daniels [43]).
- Категория: Система Duffy
Антигены Fya и Fyb появляются на 6-7-й неделе эмбрионального развития (Toivanen и соавт. [158, 159]), выраженность их такая же, как на эритроцитах взрослых, и остается неизменной на протяжении всего периода внутриутробного развития плода.
Противоречивые результаты получены при изучении сроков появления антигена Fy6. В одних исследованиях появление указанного лиганда констатировалось в одинаковые сроки с появлением гликопротеинов Lutheran, т. е. на 2-3-й мес. внутриутробного развития, в других - приблизительно в одно и то же время с формированием гликофоринов С (Bony и соавт. [20], Daniels, Green [44], Southcott и соавт. [151]).
Эритроциты Fy(a+b-) и Fy(a-b+) несут по 13 000-14000 антигенных участков Fya и Fyb на одну клетку. Количество антигенных участков Fya и Fyb на эритроцитах Fy(a+b+) вдвое меньше - по 6000-7000 (Masouredis и соавт. [103]). После обработки эритроцитов папаином число антигенных участков редуцируется более чем на 85 %.
Nichols и соавт. [124], Riwom и соавт. [140] с помощью радиоиммунного метода, используя моноклональные антитела анти-Руб, обнаружили на одном эритроците от 6000 до 12 200 участков антигена Fy6.
Уровень экспрессии антигена Fy6 на 50 % выше на ретикулоцитах, чем на зрелых эритроцитах (Woolley и соавт. [ 181,182]).
Помимо эритроцитов антиген Fy6 выявлен на клетках эндотелия посткапиллярных венул во всех органах, за исключением печени (Chaudhuri и соавт. [27], Hadley и соавт. [61]), а также на волокнах Пуркинье нейронов (Horuk и соавт. [72]). Анализ кодирующих последовательностей ДНК подтвердил, что почечные и эритроидные изоформы Duffy-гликопротеинов практически идентичны. Небольшие различия в их мол. массе, вероятно, обусловлены неодинаковым гликозилированием (Chaudhuri и соавт. [27], Hadley [62], Neote и соавт. [123]).
Dufly-гликопротеины выявлены с помощью кроличьих антител в эпителии почечных канальцев и альвеол (Chaudhuri и соавт. [27]), а также в неэритроидных тканях негров Fy(a-b-) (Peiper [132]).
РНК-транскрипты гена FY были выявлены методом гибридизации в костном мозге лиц фенотипов Fy(a+b-), Fy(a+b) и Fy(a~b+), но не Fy(a-b-) (Chaudhuri и соавт. [28, 29]). Эти транскрипты обнаруживали в тканях легких, мышц, селезенки, толстой кишки, сердца, поджелудочной железы, почек и головного мозга (Chaudhuri и соавт. [29],
Hadley и соавт. [61], Neote и соавт. [123]), а также в тканях органов негроидов с фенотипом Fy(a-b-) (Chaudhuri и соавт. [29]). Хотя величина большинства транскриптов составляла 1,35 кб, Le Van Kim и соавт. [86] установили, что в тканях головного мозга преобладали транскрипты величиной 7,5 кб. Два вида транскриптов различались между собой в области нетранслируемого 5'-региона, однако кодировали синтез одного и того же пептида. Тем не менее, Neote и соавт. [123] сообщили, что в головном мозге плода транскрипты имели величину 8,5 кб, а у взрослых-размером 1,35 кб.
Антигены Fyan Fyb не выявлены на лимфоцитах, моноцитах, нейтрофилах и тромбоцитах (Dunstan и соавт. [50, 51]).
- Категория: Система Duffy
Fy3
Антиген Fy3 присутствует на эритроцитах всех людей, за исключением Fy(a-b-). Он широко распространен среди европеоидов и монголоидов, но встречается редко среди жителей некоторых регионов Западной Африки. В отличие от факторов Fya и Fyb, антиген Fy3 устойчив к действию протеолитических ферментов (Albrey et al [10], Buchanan и соавт. [23], Daniels [42], Mannessier и соавт. [100], Oberdorfer и соавт. [126]). Эритроциты приматов содержат антиген Fy3, а антигены Fya и Fyb на них отсутствуют (Tippett, Gavin [157])
Антитела анти-БуЗ впервые обнаружены Albrey и соавт. [10] у австралийки Fy(a-b-), имевшей 3 беременности и получавшей гемотрансфузии. Антитела реагировали одинаково интенсивно с эритроцитами Fy(a+b-), Fy(a+b+) и Fy(a~b+), не разделялись на анти-Руа и анти-Fy15 дифференциальной адсорбцией. Четыре других найденных образца антител анти-РуЗ реагировали несколько интенсивнее с эритроцитами, содержавшими антиген Fya (Mannessier и соавт. [100]).
Антитела анти-РуЗ редко вырабатываются у негроидов, хотя имеются сообщения о выявлении у них нескольких таких образцов (Jensen и соавт. [80], Kosinski и соавт. [85], Molthan, Crawford [114], Oakes и соавт. [125], Oberdorfer и соавт. [126], Sosler и соавт. [150]). Гораздо чаще у негроидов Fy(a-b-), получавших многократные гемотрансфузии, выявляли антитела aHTH-Fya. Тем не менее, антитела Duffy встречаются редко и среди этой категории реципиентов. Так, скрининг сывороток 566 реципиентов-негроидов Fy(a-b-) во Франции ни в одном случае не выявил антител системы Duffy (LePennec и соавт. [89]).
Негроиды Fy(a-b-) гомозиготны по аллелю Fy и мутации в GATA-1 для эритроидных клеток. И хотя их эритроциты имеют фенотип Fy:-3, другие клетки экспрессируют гликопротеины Duffy (Peiper и соавт. [132]). Независимо от мутации в GATA-1 структура генов Fyb и Fy гомологична, поэтому гомозиготы Fy/Fy экспрессируют антигены Fyb и Fy3 в неэритроидных тканях. Это позволяет объяснить редкость антител анти-РуЗ, а также анти-Fyb среди негроидов Fy(a-b-). В то же время они способны вырабатывать антитела анти-Fy3. Молекулярная структура фенотипа Fy(a-b-) негроидов, выработавших антитела анти-РуЗ, отличается от таковой у негроидов Fy/Fy, не содержащих указанных антител. Более того, антитела анти-Fy3, образовавшиеся у негроидов Fy(a-b-) респондеров, способны улавливать различия в Duffy-гликопротеинах, экспрессированных на разных образцах эритроцитов и неэритроидных клеток.
Антитела анти-Fy3, полученные от негроидов, реагируют слабо или вовсе не реагируют с эритроцитами новорожденных (Kosinski и соавт. [85], Molthan, Crawford [114], Oakes и соавт. [125]), а антитела той же специфичности от лиц других рас реагируют с эритроцитами взрослых и новорожденных одинаково интенсивно (Buchanan и соавт. [23], Mannessier и соавт. [100]). Антигены Duffy присутствуют на эндотелиальных клетках.
Описаны немедленные и отсроченные гемолитические трансфузионные реакции (Mannessier и соавт. [100], Olteanu и соавт. [129], Vengelen-Tyler [168]), а также случаи легкой ГБН (Kosinski и соавт. [85], Molthan, Crawford [114], Oakes и соавт. [125]), обусловленные анти-РуЗ-антителами. ГБН купировали фототерапией (Albrey и соавт. [10], Buchanan и соавт. [23]).
Мышиные моноклональные антитела анти-РуЗ в серологических реакциях проявляли себя так же, как и поликлональные аллогенные.
Исследования клеток насекомых, подвергнутых трансфекции кДНК, кодирующей гибридные молекулы Fy-гликопротеина и интерлейкина 8, показали, что моноклональные антитела анти-Fy3 распознают эпитопы, расположенные на 3-й экстрацеллюлярной петле Fy-гликопротеина (Lu и соавт. [96]). Аллогенные поликлональные антитела анти-РуЗ выявляют эпитопы более чем одного региона молекулы Fy-гликопротеина.
МКА Анти-РуЗ часто используют для быстрого поиска доноров Fy(a-b-) среди доноров негров. Потребность в многократных трансфузиях таких эритроцитов весьма велика при лечении негров, страдающих серповидно-клеточной анемией (Sandler и соавт. [143]).
Fy4
После того как Sanger и соавт. [144] обосновали возможность существования молчащего аллеля Fy, обусловливающего высокую частоту фенотипа Fy(a-b-) среди негроидов, были предприняты безуспешные попытки найти антитела анти-Fy или aHra-Fyc у реципиентов белой расы, получавших трансфузии крови от доноров негров. В 1973 г. Bezhad и соавт. [19] описали антитела, найденные у негритянки Fy(a+b+), страдавшей серповидно-клеточной анемией и получившей множество трансфузий. По характеру реагирования антитела приближались к предсказанным анти-Рус-антителам, но это не были антитела анти-Рус-специфичности. Они были обозначены анти-Ру4. Реакция антител усиливалась после обработки эритроцитов папаином [19].
Исследования, проведенные с анти-Ру4-антителами в 3 лабораториях, внесли сомнения относительно принадлежности анти-Ру4-антител к системе Duffy В частности эритроциты, лишенные Fy-гликопротеина, давали с анти-Ру4-антителами слабоположительные реакции (Behzad и соавт. [19], Buchanan и соавт. [23]). Не удалось провести генетических посемейных исследований. Единственный образец антител анти-Ру4 оказался нестабильным при хранении, в связи с чем исследования в этом направлении стали невозможными (Reid, Lomas-Francis [138]).
Fy5
Антиген Fy5 близок по серологическим свойствам антигену Fy3, но отличается от последнего тем, что отсутствует у лиц Rhnu]1 (Daniels [43], Issitt, Anstee [76], Reid, Lomas-Francis [138], Westhoff, Reid [178]). Он слабо экспрессирован на эритроцитах -D-/-D-, однако присутствует на эритроцитах лиц Fy(a-b-), не относящихся к негроидам (табл. 10.5). Подобно антигену Fy3 антиген Fy5 устойчив к действию протеолитических ферментов и присутствует в одинаковой степени на эритроцитах взрослых и новорожденных (Colledge и соавт. [36], DiNapoli и соавт. [49]).
Характер реагирования анти-Ру5-антител позволяет полагать, что они взаимодействуют со структурами, интегрированными с Rh-протеинами и Rh-ассоциированными гликопротеинами (RhAG).
Таблица 10.5
Характер реагирования антител анти-РуЗ, анти-FyS и анти-Руб
|
Эритроциты |
Реакция с антителами |
||
|
анти-РуЗ |
анти-Ру5 |
анти-Руб |
|
|
Fy(a+b-) |
+ |
+ |
+ |
|
Fy(a+b+) |
+ |
+ |
+ |
|
Fy(a-b+) |
|
+ |
+ |
|
Fy(a-b-) негров |
— |
— |
— |
|
Fy(a-b-) европейцев |
— |
+ |
нд |
|
Fy(a-b+w), (Fyx/Fyx) |
сл |
сл |
сл |
|
Rh .. null |
+ |
— |
+ |
|
-D- |
1 |
СЛ |
+ |
|
Энзимированные |
+ |
+ |
— |
« + » — положительная, « - » - отрицательная, сл - слабовыраженная реакция, нд - нет данных.
Найдено по меньшей мере 6 образцов антител указанной специфичности -все у реципиентов черной расы с фенотипом Fy(a-b-), главным образом у больных серповидно-клеточной анемией, получавших гемотрансфузии (Bowen и соавт. [21], Chan-Shu [26], Colledge и соавт. [36], DiNapoli и соавт. [49], Vengelen-Tyler [168]). Во всех случаях сыворотки сенсибилизированных содержали смесь антител aHTH-Fy5 и анти-Fyа (Vengelen-Tyler [168]). Антитела анти-Fy5 не удается разделить методами адсорбции. Эритроциты некоторых лиц с парциальными антигенами Rh слабо реагировали с анти-Ру5-антителами, еще раз подтверждая положение о том, что антигены Duffy имеют некоторую связь с антигенами резус (Meredith [104]). Эритроциты лиц, гомозиготных по гену Fy\ также несут слабый антиген Fy5 (Marsh [101]).
Антитела анти-Ру5 описаны как причина замедленных гемолитических посттрансфузионных реакций (Bowen [21], Chan-Shu [26], Vengelen-Tyler [168]). У одного больного описаны 2 трансфузионные реакции, одна из которых была обусловлена антителами анти-Fy3, другая - анти-Fy5 (Bowen [21]).
Nichols и соавт. [124], Riwom и соавт. [140] получили 2 образца мышиных моноклональных антител против часто встречающегося антигена, напоминавшего Fy3. Однако в отличие от антигена Fy3, открываемый полученными антителами антиген разрушался после обработки папаином, фицином и химотрипсином. Антиген получил обозначение Fy6, соответствующие антитела - анти-Руб.
Антитела анти-Руб аллогенного происхождения не найдены (Daniels [43]).
Оба образца упомянутых моноклональных антител распознавали линейные эпитопы, представленные аминокислотными остатками QLDFEDV в позициях 19-25 на N-терминальном экстрацеллюлярном домене Fy-гликопротеина (Wasniowska и соавт. [170,173, 174]). Удалось выделить Fy-гликопротеин, напоминавший структуру, делающую возможной инвазию эритроцитов малярийными паразитами Plasmodium vivax (Chaudhuri и соавт. [30], Riwom и соавт. [140]).
Антиген Fy6 присутствует на эритроцитах низших обезьян.
- Категория: Система Duffy
Установление нуклеотидных замен, обусловливающих специфичность Fya, Fyb и Fy, позволило проводить генотипирование плодов беременных, сенсибилизированных к антигену Fya, с целью оценки риска ГБН (Goodrick и соавт. [55]). Нуклеотидная замена G 159 А, определяющая различия Fya/Fyb, создает сайт рестрикции Вап\ в аллеле Fya, а замена Т 67 С - сайт Styl, определяющий различия Fy/Fyab аллеля Fy (Iwamoto и соавт. [78], Tournamille и соавт. [160]). Генотип устанавливают с помощью ПЦР с использованием аллельспецифиче-ских праймеров С 67 (Fy), Т 67 (Fya или Fyb), G 159 (Fya), А 159 br или Fy) jpc. 1u.4j (riessner и соавт. [69], Mullighan и соавт. [119], Olsson и соавт. [127]) Использование комбинаций пар праймеров позволяет усилить продукты реакции и проследить последовательности кодонов (см. табл. 10.4).

Больные серповидно-клеточной анемией, систематически получающие гемотрансфузии, часто образуют антитела к различным аллоантигенам эритроцитов, в том числе антигенам Duffy. Генотипирование доноров и реципиентов облегчает процесс подбора совместимых пар по указанной системе. Люди Fy(a+b-) отличаются по способности вырабатывать клинически значимые анти-Руь-антитела, которые могут вырабатываться только у лиц с генотипом Fya/Fya, но не Fya/Fy, поскольку аллель Fy кодирует синтез Руь-гликопротеина в неэри-троидных клетках. Таким образом, иммунная система лиц Fya/Fy не воспринимает антиген Fyb как чужеродный.
- Категория: Система Duffy
Moore и соавт. [115], используя методы иммунопреципитации, установили, что антиген Fya присутствует на 3 протеинах, имеющих мол. массу 39, 64 и 85 кДа. Иммуноблоттинг с антителами анти-Fy* позволил выявить структуры с мол. массой 35 - 43 (Hadley и соавт. [60]) и 40-50 кДа (Tanner и соавт. [156]). Субстрат, выделенный электроэлюцией из протеинов 35-43 кДа, ингиби-ровал антитела анти-Fy3 [60]. Аналогичные результаты получили Nichols и соавт. [124], Riwom и соавт. [140], использовавшие иммуноблоттинг с монокло-нальными антителами анти-Руб.
Обработка эритроцитов эндо-Р-гликаназой, N-гликаназой и сиалидазой снижала мол. массу выделяемых с помощью иммуноблоттинга Fy-гликопротеинов до 4 кДа (Hadley и соавт. [60], Tanner и соавт. [156]).
Riwom и соавт. [140] и Wasniowska и соавт. [172] выделили низкомолекулярные фракции из очищенных Fy-гликопротеинов и третичных структур, полученных из них. Указанные исследования свидетельствовали о том, что Fy-гликопротеины N-гликозилированны и слабо О-гликозилированы. Разная степень N-гликозилирования, вероятно, и обусловливает колебания мол. массы Fy-гликопротеинов в широком диапазоне (Chaudhuri и соавт. [30]).
Chaudhury и соавт. [30] посредством иммунопреципитации моноклональны-ми антителами анти-Руб выделили Fy-гликопротеин с мол. массой 36-43 кДа вместе с олигомерами большей мол. массы, последние также реагировали с указанными антителами. Другие из сопутствующих протеинов не реагировали в реакции иммунного окрашивания с антителами анти-Руб. Картирование пептидов, выделенных из эритроцитов Fy(a+b-) и Fy(a-b+), существенных различий не выявило (рис. 10.2).

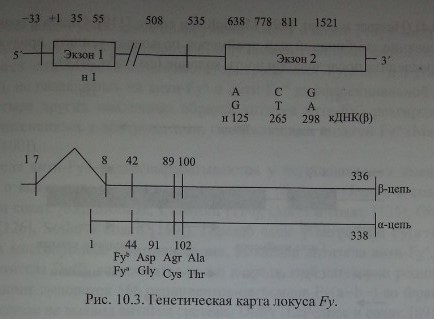
Chaudhury и соавт. [28], изучив последовательность аминокислот в очищенных Duffy-гликопротеинах, сконструировали олигонуклеотидные прай-меры и с помощью ПЦР изменили кодирующий участок ДНК, полученной от лиц Fy(a-b+). Затем этот продукт был использован для выделения кодирующей ДНК из библиотеки ДНК костномозговых клеток человека. В результате исследования получен пептид из 338 аминокислот с мол. массой 35,7 кДа (рис. 10.3).
Neote и соавт. [123] установили, что Fy-гликопротеин структурирован в виде а-спиралей, 7 раз пересекающих мембрану эритроцитов, имеет экстрацеллюлярный N- и цитоплазматический С-домены (см. рис. 10.1). Такое строение свойственно хемокиновым рецепторам (Л и соавт. [81], Murdoch, Finn [120]). Экстрацеллюлярный домен, состоящий из 65 аминокислот, имеет 3 участка N-гликозилирования - позиции 16, 27 и 33. Антитела анти-Руб реагировали с синтетическим пептидом, представляющим собой фрагмент N-терминального домена Fy-гликопротеина.
Первая из полученных образцов Duffy кДНК была представлена одним эк-зоном (Chaudhuri и соавт. [29], Iwamoto и соавт. [79], Tournamille и соавт. [160]).
Позднее Iwamoto и соавт. [77] показали, что в неэритроидных тканях преобладают транскрипты, состоящие из двух экзонов, разделенных 479 парами кодонов. Первый экзон кодирует семь N-терминальных аминокислот Fy-гликопротеина, включая инициирующий трансляцию метиониновый кодон. Терминальный участок молекул Fy-гликопротеина представлен последовательностью MGNCLHRAEL (Iwamoto и соавт. [77]).
В регионе 5! гена FY нет ТАГА- и СААТ-боксов, однако имеется несколько участков SP1 и GATA, препятствующих транскрипции (Iwamoto и соавт. [79], Tournamille и соавт. [160]).
- Категория: Система Duffy
Фенотип Fy(a-b-) был впервые выявлен Sanger и соавт. [144] при исследовании крови доноров (американских негров) сыворотками aHra-Fya и анти-Fyb. Оказалось, что большинство лиц указанной расовой принадлежности являются Fy(a-b-). Такие результаты позволили высказать предположение о существовании рецессивного аллеля в локусе Duffy. Этот молчащий аллель получил обозначение Fy. Частота фенотипа Fy(a-b-) составила 63 % среди негров Нью-Йорка, Вест-Индии (Race, Sanger [136]) и Южной Африки (Olsson и соавт. [127]), а среди других африканских популяций она оказалась еще более высокой (Mourant и соавт. [118]). Все 1168 обследованных жителей сельских районов Гамбии имели фенотип Fy(a-b-) (Welch и соавт. [176]).
Данные посемейных исследований в 53 негритянских семьях позволили подтвердить первоначальное предположение о существовании молчащего аллеля Fy (Race, Sanger [136]). Эффект дозы, выявляемый некоторыми образцами антител анти-Fy3, позволил установить, что практически все европеоиды Fy(a+b-) имеют генотип Fya/Fya и содержат двойную дозу антигена Fya, а негроиды имеют генотип Fya/Fy, их эритроциты несут 1 дозу антигена Fya и реагируют с антителами анти-Fy3 менее интенсивно, чем эритроциты Fy(a+) европеоидов (Sanger и соавт. [144]).
Хотя эритроциты Fy(a-b-) лишены ^^-гликопротеина (Chaudhuri и соавт. [28, 30]), последний у негроидов экспрессирован на клетках эндотелия посткапиллярных венул в мягких тканях и синусах селезенки (Peiper и соавт. [132]). Транскрипты гена F7 (иРНК) не были выявлены в костном мозге лиц Fy(a-b-), однако их обнаружили в других тканях (легкие, селезенка, толстая кишка) (Chaudhuri и соавт. [29]). Кодирующая последовательность аллеля Fy оказалась идентичной кодирующей последовательности аллеля Fyb (Chaudhuri и соавт. [28, 29], Iwamoto и соавт. [79], Mallinson и соавт. [99], Peiper и соавт. [132], Tournamille и соавт. [164]), однако в промоторной области имелась нуклеотидная замена Т > С, расположенная на 33 позиции выше стартовой точки считывания для эритроидных транскриптов, на 66 позиций выше такой же точки для образования большого транслирующего кодона (позиция 67), включая участок рестрикции Styl (Iwamoto и соавт. [78], Tournamille и соавт. [160]). Эта мутация внутри последовательности GATA (с заменой СТТАТСТ на СТТА£СТ) прерывает связывание эритроидспецифического фактора транскрипции GATA-1 и тем самым препятствует экспрессии антигенов FY на эритроидных клетках, при этом другие типы клеток не затрагиваются.
Iwamoto и соавт. [78], Tournamille и соавт. [160] провели эксперименты по трансфекции эритроидной клеточной линии человека (HELa) и линии эн-дотелиальных клеток промоторным регионом гена Fybn гена, кодирующего хлорамфениколацетилтрансферазу (ХАТ). В клетках, подвергнутых трансфекции, отметили высокий уровень ХАТ-активности. Трансфекция клеток фрагментами ДНК, содержавшими Fy-специфическую Т 33 С GATA-мутацию, не приводила к появлению ХАТ-активности в эритроидных клетках, однако в клетках эндотелиальной линии ее наблюдали.
Из 1062 обследованных жителей Папуа - Новой Гвинеи 23 оказались гетерозиготными по /^-специфической последовательности Gly 42 и GATA-мутации, типичной для аллеля Fy. Частота аллеля составила 0,022 (Zimmerman и соавт. [184]). По-видимому, указанный аллель ведет себя как молчащий в отношении эритроидных клеток, поскольку эритроциты таких лиц имеют слабую экспрессию антигена Fy6 по сравнению с эритроцитами гомозигот Fya/Fya.
Фенотип Fy(a-b-), типичный для негроидов, встречается крайне редко среди представителей других рас и этнических групп (Mourant и соавт. [118]). Он не был найден среди 6 тыс. белых жителей Австралии при их обследовании с помощью сыворотки aHTH-Fy3 (Albrey и соавт. [10]).
Chown и соавт. [35], Lewis и соавт. [92] рассчитали, что частота гена Fy среди лиц белой расы составляет около 0,001; ожидаемая частота фенотипа Fy(a-b-) - 1 на 1 млн. В нескольких семьях выявлено необычное наследование генов Fya и Fyb, обусловленное, как полагают Chown и соавт. [35], присутствием аллеля Fyx.
Людей Fy(a-b-) среди европеоидов и монголоидов обычно идентифицировали в связи с обнаружением в сыворотках их крови активных антител анти-Fy3. У одной из австралийских женщин Fy(a-b-) имелась делеция 14 пар кодонов гена FY (Albrey и соавт. [10]), которая изменяла рамку считывания и приводила к прекращению трансляции (Mallinson и соавт. [99]). Молекулярно-генетические методы исследования позволили выявить у 3 неродственных лиц гомозиготность по нонсенс-мутациям, приведшим к формированию фенотипа Fy(a-b-):
- G 408 А (Trp 136 Stop) в гене Fya у англичанки (Rios и соавт. [139]);
- G 407 А (Trp 96 Stop) в гене Fyby ливанской еврейки (Rios и соавт. [139]);
- G 287 А (Trp 136 Stop) в гене Fyay индианки из Канады (Buchanan [23], Rios и соавт. [139]).
Tsuneyama и соавт. [165] наблюдали японку Fy(a-b-), без антител, имевшую делецию С327, которая приводила к остановке считывания 12 кодонов ниже 120-го.
Каждая из мутаций, описанных выше, проявляла себя отсутствием экспрессии антигенов Duffy на эритроцитах в отличие от GATA-мутаций, способствующих формированию африканского варианта фенотипа Fy(a-b-), при котором Duffy-гликопротеины экспрессированы на неэритроидных клетках.
Фенотип Fy(a-b-) без антител анти-Fy 3 описан у чешских цыганок (Libich и соавт. [94], Pisacka и соавт. [134]), белой женщины шотландско-швейцарского происхождения, имевшей GATA-мутацию, характерную для негроидов.
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}