Поиск по сайту
- Вы здесь:
-
Главная

-
book
-
Группы крови человека
- Система Cartwright и система Xg
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...

- Категория: Система Cartwright и система Xg
Herron и Smith [53] посредством иммунопреципитации субстрата антителами анти-Xg3 установили, что антигенные эпитопы Xga эритроцитарной мембраны относятся к сиалогликопротеинам. Установлена мол. масса вещества (22,5-28 кДа). Обработка эритроцитов сиалидазой приводила к уменьшению мол. массы. Гликопротеин, определяющий специфичность Xg, тесно связан с белком CD99 (Petty, Tippett [71 ], Fouchet и соавт.
Кодируемый геном XG протеин состоит из 180 аминокислот. Экстрацеллюлярный N-терминальный домен включает 142 аминокислоты, содержит 16 участков О-гликозилирования, но не имеет участков N-гликозилирования. Остальная часть Xg-протеина представлена трансмембранным и црггоплазматическим С-терминальным доменами (рис. 14.1) (Ellis и соавт. [32]).
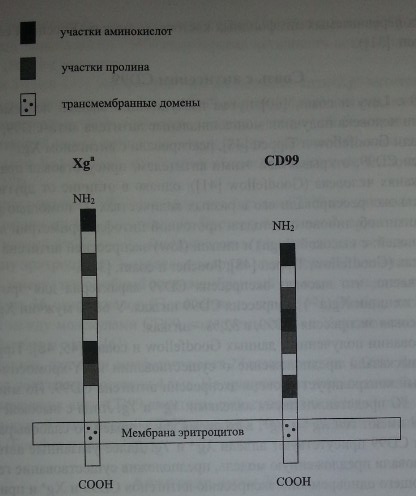
Строение гликопротеинов Xga и CD99
Строение гликопротеинов Xga и CD99
Транскрипты гена XG, помимо эритроидных клеток и фибробластов, выявлены в скелетных мышцах, сердце, плаценте, клетках предстательной и щитовидной желез, позвоночнике и трахее (Fouchet и соавт. [37]), Некоторое количество транскриптов XG обнаружено в легких, почках, яичке, а также в тканях плода и некоторых перевиваемых лимфоидных клеточных линиях.
- Категория: Система Cartwright и система Xg
Обследование 6784 жителей стран Северной Европы показало (табл. 14.2), что среди женщин частота фенотипа Xg(a+) составляет 89 %, в то время как среди мужчин - 66 % (Sanger и соавт. [80], Daniels [19], Haldane [51]).
Распределение антигена Xg у мужчин и женщин
|
Показатель |
Частота, % |
||
|
у мужчин |
у женщин |
||
|
Фенотип |
Xg(a+) |
65,9 |
88,7 |
|
Xg(a-) |
34,1 |
11,3 |
|
|
Генотип |
Xga/Xga |
43,4 |
|
|
XgVXg |
65,9 |
45,0 |
|
|
Xg/Xg |
34,1 |
11,6 |
|
Антиген Xga обнаружен у представителей всех исследованных популяций. Наибольшая частота гена Xgaконстатирована у аборигенов Новой Гвинеи и Австралии, жителей о. Сардинии и бразильцев ген Xga не вполне развит к моменту рождения. Его экспрессия на эритроцитах новорожденных ниже, чем на эритроцитах взрослых (Мауг [65], Toivanen, Hirvonen [94]). Вещество Xga формируется в относительно поздние сроки внутриутробного развития. Из 54 плодов от 6 до 20 недель развития только 19 были Xg(a+), что существенно ниже частоты встречаемости этого антигена в популяции (Toivanen, Hirvonen [93]). Антиген Xg(a+) появляется у плодов начиная с 12 недель развития. У 5-10 % мальчиков Xg(a+) при рождении указанный антиген не удается выявить серологическими методами (Szabo и соавт. [90]). По мере взросления организма выраженность его на эритроцитах возрастает (Сатрапа и соавт. [15]).
В процессе эритропоэза in vitro антиген Xga появлялся на клетках после гликофорина А и протеина полосы 3, но раньше Rh-протеина (Daniels, Green [20]).
Антиген Xga не является сильным иммуногеном. На эритроцитах гетерозиготных мужчин и гомозиготных женщин он выражен одинаково. У женщин, гетерозиготных по гену Xga, экспрессия антигена может быть снижена. До 10 % гетерозиготных женщин имеют слабовыраженный антиген Xga (Race, Sanger [73]). Слабые варианты Xga среди мужчин редки.
Количество антигенных участков Xga на эритроците, по данным разных авторов, варьирует от 159 (Foucher и соавт. [36]) до 9000 (Szabo и соавт. [90]).
В 1974 г. Fellous и соавт. [34] обнаружили антиген Xga на фибробластах и гибридомных клетках человек - мышь. Антиген выявлялся одновременно с другими продуктами Х-ассоциированных локусов. Тем самым было еще раз подтверждено, что антиген Xga контролируется геном, расположенным на Х-хромосоме. При инактивации Х-хромосомы антиген Xga на клетках человека не выявлялся (Hsu и соавт. [54]).
Антиген Xga разрушается бромелином, фицином, папаином, проназой, трипсином и химотрипсином (Habibi и соавт. [49], Herron, Smith [53]), однако устойчив к воздействию сиалидазой.
Частота генов XG у разных народов
|
Популяция |
Количество обследованных |
xga1 |
Xg |
Источник |
|
Аборигены Новой Гвинеи (папуасы) |
263 |
0,85 |
0,15 |
[85] |
|
Австралийские аборигены |
352 £ |
0,79 |
,0,21 |
[85] |
|
Индейцы навахо |
308 |
0,77 |
0,23 |
[28] |
|
Жители о. Сардиния |
322 |
0,76 |
0,24 |
[89] |
|
Бразильцы белые |
1 078 |
0,74 |
0,26 |
[68] |
|
Бразильцы мулаты |
786 |
0,62 |
0,38 |
[68] |
|
Бразильцы негры |
827 |
0,57 |
0,43 |
[68] |
|
Евреи Израиля, сефарды |
201 |
0,68 |
0,32 |
[1] |
|
Японцы |
529 |
0,68 |
0,32 |
[67] |
|
Жители стран Северной Европы |
15 716 |
0,66 |
0,34 |
[16,27,52,65,66, 80] |
|
Индусы, Бомбей |
100 |
0,65 |
0,35 |
[10] |
|
Индусы, Сингапур |
|
0,57 |
0,43 |
[77] |
|
Китайцы, континентальный Китай |
171 |
0,60 |
0,40 |
[28] |
|
Китайцы, Тайвань |
178 |
0,53 |
0,47 |
[28] |
|
Китайцы, Гонконг |
1300 |
0,49 |
0,51 |
[63] |
|
Китайцы, Сингапур |
165 |
0,45 |
0,55 |
[11,77] |
|
Испанцы |
636 |
0,59 |
0,41 |
[96] |
|
Таицы |
181 |
0,57 |
0,43 |
[74] |
|
Греки |
638 |
0,55 |
0,45 |
[38] |
|
Негры, Нью-Йорк, жители Ямайки |
219 |
0,55 |
0,45 |
|
|
Малайцы, Сингапур |
72 |
0,54 |
0,46 |
[77] |
|
Тайваньцы (аборигены) |
164 |
0,38 |
0,62 |
[28] |
- Категория: Система Cartwright и система Xg
Хромосомы, определяющие пол, так же как и аутосомы, принимают участие в формировании групповых признаков крови человека. В 1962 г. были открыты группы крови, которые передаются по наследству с Х-хромосомой.
Mann и соавт. [64] выявили в сыворотке крови мужчины белой расы (мистера And) необычные антитела, которые в отличие от всех известных ранее существенно чаще реагировали с эритроцитами женщин, чем мужчин. Посемейные исследования подтвердили, что антиген, получивший обозначение Xga, находится под контролем гена Xga (XG1\ сцепленного с Х-хромосомой. Поскольку антиген, антитетичный антигену Xga, не найден, было сделано заключение, что систему Xg образуют два гена: продуктивный ген-Xga(XGl) и его молчащий аллель-Xg (XG0).
С группами крови Xg тесно связан антиген CD99, находящийся под контролем гена МС2, присутствующего как на Х-, так и на Y-хромосоме (Goodfellow, Tippett [48]). В связи с этим антигены Xgaи CD99 оказались весьма ценными маркерами при изучении сцепленности различных признаков с полом, X-Y-гибридизации и рекомбинации, инактивации Х-хромосомы, Х- и Y-анэуплоидии, патологии формирования пола, в частности мужского типа у лиц XX.
Антиген Xga
Наследование
Пол человека детерминирован двумя хромосомами: XX у женщин, XY у мужчин. Ген Xga (XG1) расположен на Х-хромосоме, на Y-хромосоме он отсутствует. Если на Х-хромосоме мужчины присутствует аллель Xga, то он имеет фенотип Xg(a+). Если на Х-хромосоме мужчины отсутствует аллель Xga, а имеется молчащий ген Xg9 то формируется фенотип Xg(a-). Женщины Xg(a+) могут быть гомозиготными (Xga/Xga) или гетерозиготными (Xga/Xg\ в то время как все мужчины Xg(a+) гетерозиготны (Xga/Xg). Соответственно антиген Xga чаще встречается у женщин.
Анализ распределения групп крови Xg в 2540 семьях с 5824 детьми в Северной Европе (Sanger и соавт. [80]), а также во многих других семьх, включая жителей Канады (Chown и соавт. [16]), Сардинии (Suniscalo и соавт. [89]), Израиля (Adam и соавт. [1]) и Японии (Nakajima и соавт. [67]), подтвердил, что антиген Xga передается по наследству как Х-сцепленный доминантный признак. Исключения из этого правила, т. е. передача гена Xga с Y-хромосомой или другим путем, очень редки.
Таблица 14 J
Варианты фактического наследования антигена Xg
|
Фенотипы и генотипы |
|||||||
|
родителей |
детей |
||||||
|
отца |
матери |
сына |
дочери |
||||
|
Xg(a+) |
Xga |
Xg(a+) |
Xga/Xga |
Xg(a+) |
Xga |
Xg(a+) |
Xga/Xga |
|
Xg(a+)' |
Xga |
Xg(a+) |
Xga/Xg |
Xg(a-)1 |
Xg |
Xg(a+) |
XgVXg |
|
Xg(a-)2 |
Xga |
Xg(a-) |
Xg/Xg |
Xg(a+) |
Xga |
Xg(a+)2 |
Xg/Xga |
|
Xg(a-)3 |
Xg |
Xg(a+) |
Xga/Xga |
Xg(a+) |
Xga |
Xg(a+) |
Xg/Xga |
|
|
|
|
|
Xg(a-)3 |
Xg |
Xg(a-)3 |
Xg/Xg |
|
Xg(a-) |
Xg |
Xg(a-) |
Xg/Xg |
Xg(a-) |
Xg |
Xg(a-) |
Xg/Xg |
1 Данные противоречат положению о доминантном типе наследования Xga,
2 Ген Xgf2 фенотипически себя не проявляет, но передается по наследству,
3 Ожидаемый антиген Xga отсутствует.
В некоторых семьях не наблюдали Х-сцепленного наследования (табл. 14.1). Обычно мужчины Xg(a+) наследуют ген Xg61 от матери. Однако в 16 семьях сыновья женщин Xg(a-) имели группу Xg(a+) (Sanger и соавт. [78, 80], Chown и соавт. [16], Tippett, Ellis [91], Race, Sanger [73]).
Race и Sanger [73] высказали предположение, что небольшая часть Х-хромосомы, включающая участок XG, может транслоцироваться на Y-хромосому. Далее реком-бинантная Y-хромосома передается по наследству сыновьям. Таким образом, антиген Xg в части случаев может передаваться с Y-хромосомой. Однако и такой механизм наследования не объясняет встречающиеся варианты. В одной семье у мужчины Xg(a+) мать была Xg(a-) и его сын был также Xg(a-). Описан фенотип Xg(a-) у дочерей, отцы которых были Xg(a+) (Sanger и соавт. [80], Tippett, Ellis [91]). В последнем варианте наследования нельзя исключить делецию локуса XG.
- Категория: Система Cartwright и система Xg
АХЭ играет исключительно важную роль в обеспечении работы мышечного аппарата и мозга, поэтому неудивительно, что нулевой фенотип Yt(a-b-), обусловленный молчащим геном Yt-, и делеций в локусе ACHE чрезвычайно редки. Дефект гена, который может привести к недостатку столь важного для жизни фермента, по-видимому, является летальным.
Rao и соавт. [38] наблюдали больного с транзиторным фенотипом Yt(a-b-). Ему проводили медикаментозную подготовку к операции трансплантации сердца. В серологических реакциях его эритроциты тестировались как Yt(a-b-), но при этом они адсорбировали и высвобождали при элюции два из четырех образцов анти-У^-антител. Активность АХЭ на эритроцитах составляла 10 % от нормы. Сыворотка крови содержала антитела, реагировавшие одинаково интенсивно с эритроцитами Yt(a+b-) и Yt(a~b+). Антитела получили обозначение aHTH-Ytab. Они реагировали со всеми эритроцитами за исключением собственных эритроцо-тов и эритроцитов больных ПНГ (комплементчувствительная фракция). Указанные антитела можно было отнести к трансфузионно опасным, о чем свидетельствовали результаты теста с введением эритроцитов, меченных Сг51. В случае возникновения показаний к гемотрансфузии больному могли быть перелиты только его собственные эритроциты, которые и были для него заготовлены. Через 4 мес. у больного определялся слабо выраженный антиген Yta, активность АХЭ на эритроцитах возросла до 60 % и, что интересно, эритроциты стали адсорбировать anra-Ytab-антитела, которые у него ранее образовались, однако не до полного истощения.
Некоторые исследователи обратили внимание на то, что уровень АХЭ на эритроцитах больных диабетом, ниже, чем у здоровых (Testa и соавт., 1988; Suhail, Rabini, 1900). Однако Wittaker и Telen (1994) в значительно более широком исследовании не обнаружили существенного изменения концентрации АХЭ на эритроцитах больных диабетом, а также снижения экспрессии антигена Yta на эритроцитах.
Высказано предположение, что дефицит АХЭ при диабете, отмеченный в ранних исследованиях, не является истинным и может быть обусловлен тем, что кровь для исследования у больных диабетом брали натощак до инъекции инсулина.
Отмечено, что снижение концентрации АХЭ обратно пропорционально повышению уровня глюкозы. После инъекции инсулина уровни АХЭ у больных диабетом возвращались к норме.
- Категория: Система Cartwright и система Xg
Антитела анти-Yt* и aHTH-Ytb
Несмотря на небольшое число лиц Yt(a-), способных к сенсибилизации антигеном Yta, антитела к этому антигену не являются редкостью и многократно описаны в литературе [1, 2,4, 5, 8-10,12,13,18,20,24,35]. Стандартизация 79 сывороток aHTH-Yta показала, что 57 из них были моноспецифическими, 22 содержали сопутствующие антитела другой специфичности (Eckrich и соавт. [13]).
Антиген Ytb в отличие от антигена Yta является более слабым иммуногеном. Антитела aHTH-Ytb встречаются редко и практически всегда обнаруживаются в сочетании с антителами другой специфичности (Giles и соавт. [16, 17], Ferguson и соавт. [14], Ikin и соавт. [21], Wurzel, Haesler [49], Levy и соавт. [27]). Так, согласно сводке Issitt и Anstee [22], в одной из сывороток aHTH-Ytb присутствовали анти-Fyb и анти-Вра (Bishop системы Diego), в другой - анти-С и анти-Hov (DI9 системы Diego), в третьей - анти-е, в четвертой - анти-К, в пятой -анти-Е и анти-Le3.
Следует отметить, что антитела к антигенам Yta и Ytb находили у беременных или реципиентов. Они ни разу не описаны как антитела естественного происхождения. Практически все образцы антител Yt были IgG с оптимумом реагирования при температуре 37 °С в непрямой антиглобулиновой пробе. Большинство образцов относилось к субклассу IgGl, нередко в сочетании с IgG4. Некоторые образцы были представлены только субклассом IgG4. Ни в одном из случаев антитела не относились к субклассу IgG3 (Vengelen-Tyler, Morel [48], Pierse и соавт. [37]). Некоторые образцы антител анти-Yt3 обладали способностью связывать комплемент (Bergvalds и соавт. [4]), в то время как у других такая способность отсутствовала (Gobel и соавт. [18], Ballas, Sherwood [2]).
Антитела системы Yt не описаны ни разу в качестве причины ГБН, хотя в ряде случаев были выявлены у женщин Yt(a-), родивших детей Yt(a+) (Wurzel, Haesler [50], Gobel и соавт. [18], Bergvalds и соавт. [4], Bettigole и соавт. [5], Lavallee и соавт. [24], Davey, Simkins [9]). У одной женщины Yt(b-), имевшей анти-У^-антитела, родился ребенок Yt(b+) без признаков ГБН (Ferguson и соавт. [14]). Антитела системы Yt считаются клинически значимыми в практике трансфузиологии. Антитела анти-Yt3 вызвали гемолитическую посттрансфузи-онную реакцию с летальным исходом у больного серповидно-клеточной анемией (Reed и соавт. [39]). Описаны посттрансфузионные реакции немедленного типа, обусловленные антителами анти-Yt3 (Hadley и соавт. [19]). Вместе с тем у многих реципиентов, имевших анти-У^-антитела и получавших трансфузии эритроцитов Yt(a+), признаков несовместимости не наблюдалось (Dobbs и соавт. [10], Eckrich и соавт. [13]). В одном случае антитела анти-Yt3 не вызвали посттрансфузионной реакции, несмотря на то, что в тестах in vitro проявляли гемолитические свойства, свидетельствовавшие об их трансфузионной опасности (AuBuchon и соавт. [1]). Из 18 реципиентов, имевших анти-У1а-антитела, только у 3 наблюдалось уменьшение продолжительности жизни перелитых эритроцитов (Eckrich и соавт. [13]). По данным других исследователей, длительность персистенции несовместимых эритроцитов в кровотоке реципиентов варьировала в широких пределах (Gobel и соавт. [18], Bettigole и соавт. [5], Dobbs и соавт. [10], Ballas, Sherwood [2], Davey, Simkins [9], Nance и соавт. [35], Kakaiya и соавт. [23]). Лишь в отдельных случаях имелись основания полагать, что перелитые эритроциты будут быстро элиминированы антителами из кровотока (Gobel и соавт. [18], Bettigole и соавт. [5], Ballas, Sherwood [2]). В исследованиях с использованием клеток, фагоцитирующих эритроциты, сенсибилизированные антителами, также получены противоречивые данные (Gobel и соавт. [18], AuBuchon и соавт. [1], Eckrich и соавт. [13], Levy и соавт. [27], Pierse и соавт. [37], Hadley и соавт. [19], Kakaiya и соавт. [23]). В связи с этим представляется очевидным, что каждый образец антител требует индивидуального исследования, по результатам которого можно прогнозировать степень несовместимости перелитых эритроцитов и, соответственно, безопасность и лечебную эффективность планируемой трансфузии.
При отсутствии совместимых эритроцитов Yt(a-) реципиенту, имеющему анти-У1а-антитела, переливают эритроциты Yt(a+). При этом целесообразно отобрать эритроциты, дающие менее сильную реакцию in vitro с сывороткой реципиента.
Описаны анти-У^-антитела у реципиента Yt(a+). Эти антитела реагировали с эритроцитами Yt(a+) неродственных лиц и матери реципиента, но не реагировали с эритроцитами самого реципиента и его Yt(a+) отца (Mazzi и соавт. [31]). Указанное наблюдение нельзя рассматривать как доказательство существования парциальных вариантов антигена Yta, поскольку анти-У1;а-антитела, появившиеся у реципиента на 9-й день после переливания 5 доз эритроцитов Yt(a+), через несколько месяцев исчезли.
- Категория: Система Cartwright и система Xg
Из 1568 доноров, обследованных Eaton и соавт. [12], Giles и соавт. [17] в Оксфорде, только 5 имели фенотип Yt(a-), остальные - Yt(a+). Таким образом, частота антигена Yta составила 99,8 %, гена Yta - 0,9559 (табл. 13.2). Установлено, что 8 % лиц белой расы имели фенотип Yt(b+). По результатам изучения частоты антигенов YtaH Ytb рассчитана частота генотипов: Yta/Yta(0,8966), Yta/Ytb (0,1006) и Ytb/Ytb (0,0028). Расчеты показали, что существование третьего аллеля в системе Cartwright вряд ли возможно
Распределение антигенов Yta и Ytbу различных народов
|
Популяция |
п |
Частота фенотипа, % |
Частота гена |
Источник |
||
|
Yt(a+) |
Yt(b+) |
Yta |
|
|||
|
Англичане |
2 568 |
99,8 |
— |
0,9559 |
0,0441 |
[12,17] |
|
Валлийцы (уэльсцы) |
29 802 |
99,9 |
— |
0,9761 |
0,0239 |
[15] |
|
Европейцы |
1399 |
— |
8,1 |
0,9587 |
0,0413 |
[17] |
|
Французы |
7 510 |
99,7 |
8,1 |
0,958 |
0,042 |
[42] |
|
Канадцы (белые) |
659 |
100,0 |
10,6 |
0,9469 |
0,0531 |
[28] |
|
Американские негры |
714 |
— |
8,4 |
0,9571 |
0,0429 |
[50] |
|
Евреи (израильские) |
2 549 |
98,6 |
21,3 |
0,8845 |
0,1154 |
[25] |
|
Арабы (израильские) |
85 |
97,6 |
23,5 |
0,8706 |
0,1294 |
[25] |
|
Друзы (израильские) |
77 |
97,4 |
26,0 |
0,8571 |
0,1429 |
[25] |
|
Японцы |
5 000 |
100,0 |
1,0000 |
0,0000 |
[34] |
|
Примечание: « - » - нет данных.
Обследование лиц различной национальности показало, что среди евреев, арабов и друзов Израиля антиген Ytb встречается относительно часто (табл. 13.2). В то же время этот фактор не был выявлен у японцев, жителей Таиланда, а также у инков и других коренных жителей Южной Америки (Nakajima и соавт. [34], Mourant и соавт. [33]).
Giles и соавт. [17] установили среди 1399 обследованных 113 (8,1 %) лиц Yt(b+). Salmon и соавт. [42] выявили 608 человек Yt(b+) среди 7510 французов (8,1 %). Lewis и соавт. [28] обследовали 659 канадцев, из них 589 были Yt(a+b~), 70 Yt(a+b+), не было ни одного Yt(a-b+). Wurzel и Haesler [50] протестировали эритроциты 714 американских негров и обнаружили, что 60 из них (8,4 %) были Yt(b+). Эти данные показывают, что частота Yt* и Ytb одинакова у негроидов и европеоидов.
Обращают на себя внимание две популяции, в которых частота антигенов Yta и Ytb отличается от таковой в других сравниваемых группах. Так, Nakajima и соавт. [34] протестировали 5 тыс. японских доноров, из них 70 были протестированы анти-У^-сывороткой. Все японцы были Yt(a+), ни одного индивида Yt(b+) не обнаружено. Возможно, для японцев характерен исключительно фенотип Yt(a+b-).
Отмечена более высокая, чем ожидалась, частота анти-У1:а-антител у израильтян. Когда появились тестовые анти-У1ь-реактивы, было установлено, что частота этого антигена у жителей Израиля существенно выше, чем в других географических зонах. Повышенная частота антигена Ytb в этом регионе привела к некоторому увеличению частоты фенотипа Yt(a-b+), что могло повлиять на частоту аллоиммунизации лиц Yt(a-b+). Из 1683 целенаправленно протестированных израильских евреев, арабов и друзов 26 имели фенотип Yt(a-b+). Иными словами, среди этих народов лиц Yt(a-b+) обнаруживают с частотой 1 на 65. Частота встречаемости этого фенотипа среди англичан, французов и негров составляет примерно 1 на 500, то есть почти в 10 раз меньше.
Результаты двух посемейных исследований свидетельствовали о кодоминантном наследовании генов Ytan Ytb в соответствии с законом Менделя. Данных о существовании молчащего аллеля не получено (Giles и соавт. [17], Lewis и соавт. [28]).
С антигенными различиями Yta/Ytb ассоциированы две нуклеотидные замены внутри гена ACHE. Одна из них, С 1057 А в экзоне 2, кодирует замену гисти-дина на аспарагин в положении 353. Другая мутация, С 1432 Т в экзоне 3, фе-нотипически не проявляется. Третья замена нуклеотидов, в экзоне 5, также не проявляла себя в зрелом протеине (Battels и соавт. [3]).
Исследование АХЭ электрического ската Torpedo californica, наиболее изученной формы этого фермента, показало, что лишь аминокислотная замена в позиции 353 приводит к формированию иммуногенного участка, способного стимулировать синтез специфических антител. Различия в антигенной структуре. АХЭ не влияют на ее активность (Masson и соавт. [30]).
Антиген Yta устойчив к действию трипсина, а-химотрипсин его разрушает. Обработка эритроцитов фицином и папаином дала разноречивые результаты, которые зависели от того, какими образцами анти-У^-антител проводили тестирование энзимированных эритроцитов (Eaton и соавт. [12], Vengelen-Tyler, Morel [48], Morton [32], Rouger и соавт. [41], Daniels [7]). В некоторых случаях обработка указанными протеолитическими ферментами усиливала реакцию. К действию сиалидазы антиген Yta не чувствителен (Rouger и соавт. [41], Daniels [7]).
Антигены Yta и Ytb разрушаются под воздействием дитиотрейтола, 2-ами-ноэтилизотиоурониумбромида (АЕТ), 2-меркаптоэтанола (Branch и соавт. [6], Levene, Harel [26], Shulman и соавт. [43]). Денатурация зависела от концентрации дитиотрейтола при обработке эритроцитов. Когда применяли 200 мМ препарата, как Yta, так и Ytb денатурировались. При использовании 50 и 100 мМ растворов дитиотрейтола активность антигена Yta снижалась пропорционально повышению концентрации раствора ДТТ, но не подавлялась полностью.
Shulman и соавт. [43] показали, что антиген Ytb денатурируется, когда эри-троцты обрабатывают 500 мМ раствором 2-меркаптоэтанола.
Уместно упомянуть, что применение АЕТ для создания искусственных эритроцитов Kellnull подверглось критике, поскольку этот препарат разрушает и другие, не относящиеся к системе Kell, антигены. Однако, когда Levene и Harel [26] обработали АЕТ эритроциты Yt(a+), денатурация антигена Yta не была такой полной, как денатурация антигенов системы Kell. Девять из 15 сывороток aHTH-Yta не реагировали с эритроцитами, обработанными АЕТ, другие 6 все же реагировали, хотя не так сильно, как с необработанными эритроцитами.
Антигены Yta и Ytb определяются на эритроцитах новорожденных: Ytb выражен слабо, в то время как экспрессия Yta не отличается от таковой у взрослых. Giles и соавт. [17] установили, что экспрессия антигена Ytb у новорожденных не изменена. Слабую экспрессию антигена Yta наблюдали у недоношенных (Eaton и соавт. [12], Ferguson и соавт. [14]); 8 из 10 образцов эритроцитов не реагировали с анти-У1ь-сывороткой, остальные 2 реагировали слабо (Gobel и соавт. [18]).
АХЭ присутствует в нервной и мышечной тканях и на эритроцитах (Taylor [46]). Данных о распределении этого фермента в других тканях мало. Антиген Yta не был выявлен на лимфоцитах, гранулоцитах и моноцитах при исследовании методом проточной цитофлюориметрии (Dunstan [11]).
- Категория: Система Cartwright и система Xg
Telen и соавт. [47] показали, что большинство образцов анти-У^-антител не реагирует с фракцией комплементчувствительных эритроцитов больных парок-сизмальной ночной гемоглобинурией (ПНГ), однако взаимодействует с фракцией эритроцитов, не чувствительных к комплементу. Установлено, что упомянутые комплементчувствительные эритроциты в значительной степени лишены глико-зилфосфатидилинозитола (ГФИ). У двух больных ПНГ комплементчувствительные эритроциты были Yt(b+). Эти результаты интерпретированы авторами как указание на то, что эпитопы YtaH Ytb несет ГФИ-ассоциированный гликопротеин.
В 1991 г. Spring и соавт. [45] показали, что антигены Cartwright на эритроцитах несет ГФИ-ассоциированный гликопротеин АХЭ (см. рис. 13.1). Путем иммунопреципитации антителами aHTH-Yta и анти-Yt15 выделен субстрат с мол. массой 72 кДа. По электрофоретической подвижности он был идентичен веществу, выделенному с помощью моноклональных антител к АХЭ эритроцитов. Субстрат обладал ацетилхолинэстеразной активностью, о чем свидетельствовали результаты экспериментов Petty [36] с иммобилизацией моно- и поликло-нальных антител анти-Yt3 и анти-Yt15.
АХЭ эритроцитов подвергается TV-гликозилированию. Как установили Spring и соавт. [45], обработка субстрата, выделенного посредством иммунопреципитации, N-гликаназой приводила к уменьшению его мол. массы с 72 до 63 кДа. С помощью МКА к АХЭ установлено, что один эритроцит несет 3-5 тыс. АХЭ-участков. При использовании для аналогичных исследований Fab-фрагментов указанных антител выявлено 7-10 тыс. участков связывания. Это дало основания Spring и соавт. [45] полагать, что АХЭ присутствует в мембране эритроцитов в форме димера.
Как указывалось выше, АХЭ играет важную роль в передаче нервных импульсов в исполнительные органы и ткани. Ацетилхолин обеспечивает передачу электроимпульсных сигналов от терминальных участков нервов в мышечные клетки. Затем ацетилхолин быстро разрушается АХЭ путем гидролиза для прекращения последующей передачи нейроимпульсов.
АХЭ присутствует в различных тканях в разных изоформах, обусловленных альтернативным сплайсингом гена ACHE (Taylor [46], Li и соавт. [29]). Фрагменты кДНК гена ACHE обнаружены в библиотеке генов из клеток фетальной мышечной ткани и нервной ткани взрослых лиц (Soreg и соавт. [44]). Полная кодирующая последовательность гена установлена Li и соавт. [29] при анализе космидной библиотеки генома человека.
Три экзона кодируют сигнальный пептид и N-терминальный участок, состоящий из 535 аминокислот (рис. 13.3). За счет альтернативного сплайсинга следующего экзона возникают структурные изменения в С-терминальном домене, позволяющие ГФИ присоединяться к эритроидным клеткам.
Аминокислотная последовательность протеина АХЭ
Новости медицины
Много миллионов человек в мире принимают статины, но исследования показывают, что только 55% из тех, кому рекомендуется принимать статины, принимают их. Это большая проблема, потому что исследования также показывают, что те из группы...
Связанное с беременностью высокое кровяное давление может привести к долгосрочным сердечным рискам, показывают новые исследования.
Кэролин Консия, столкнулась с более серьезными последствиями репрессий против назначения опиоидов, когда узнала, почему сын ее подруги покончил с собой в 2017 году.
Новое исследование показывает, что психические заболевания не являются фактором большинства массовых расстрелов или других видов массовых убийств.


{kind=link}