




Различают 2 типа Rh ull: регуляторный (супрессорный) и аморфный (отсутствие генетического материала).
Регуляторный тип формируется в результате взаимодействия генов RH, производящих соответствующие полипептиды, и генов Х]г, Х°г, которые управляют функцией локуса RH. Ген Х1г (доминантный) запускает нормальную продукцию Rh-полипептидов, а его редкий аллель Х°г (рецессивный) блокирует их синтез.
Ген Х]г широко распространен, поэтому большинство людей являются гомозиготами - Xlr/Xlr. При таком генотипе унаследованные от родителей гены RH функционируют нормально. При гетерозиготном варианте (Xlr/X°r) экспрессия антигенов, по данным одних авторов, не нарушена [331, 532], по данным других - снижена [353].
У людей, гомозиготных по супрессорному гену, Х°г/Х°г, присутствующие ДЯ-гены не функционируют. На рис. 4.11 представлен пример наследования ре-гуляторного типа Rh 1Г.
Аморфный тип Rhnull обусловлен отсутствием генетического материала, кодирующего продукцию Rh-полипептидов, по-видимому, вследствие делеции.
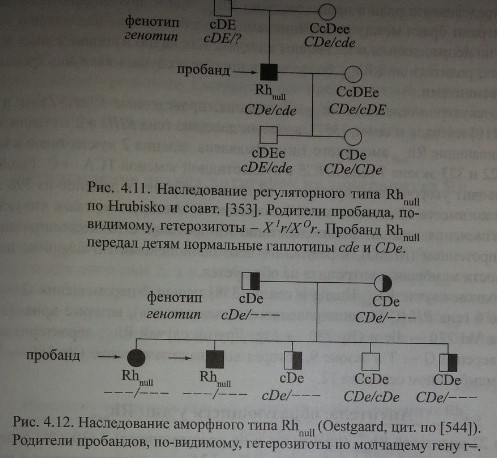
Получены данные, которые можно трактовать в пользу существования молчащего гена щ [371]. Пример наследования аморфного типа Rhnull представлен на рисунке 4.12. Родители пробандов - сибсы. Их фенотипы cDe и CDe создают видимость гомозиготности по генам с и С.
Судя по фенотипу детей, оба родителя были гетерозиготами: один, вероятно, cDe/r~> второй i CDe/r^. Двое детей имели одинаковый генотип г=/г=. Трое
других детей имели генотипы cDe/r=, CDe/r= и CDe/cDe. В приведенном примере недостает представителей третьего поколения, фенотипы которых могли бы подтвердить, что от родителей с аморфным типом Rhnul возможно рождение детей с таким же аморфным типом.
Обследование малого числа поколений в семьях не позволяет идентифицировать аморфный тип Rhnull, поскольку трудно получить убедительные доказательства того, что конкретный обследуемый человек генетически r=/r=, а не Х°г/Х°г. Генотип Х'г/Х°г также может приводить к уменьшению экспрессии антигенов Rh. Однако совокупность фактов, полученных при обследовании большого количества семей, позволила исследователям сделать вывод, что наследование Rhnull через молчащий ген г= вполне возможно.
Из 14 обладателей фенотипа Rhnull (см. табл. 4.24) у 9 был обнаружен регуля-торный тип Rhnull, у 3 - аморфный, у 2 - тип Rhnull не установлен.
Среди упомянутых выше 33 обследованных лиц Rh „ 20 имели регулятор-ный тип, 3 - аморфный тип, у остальных 10 человек генетическую основу нулевого фенотипа установить не удалось. Последующие многочисленные исследования подтвердили, что оба типа, регуляторный и аморфный, встречаются крайне редко, причем один из них, регуляторный, преобладает [151, 153, 331, 353, 532]. В некоторых случаях семейные исследования не могли выявить генетическую основу фенотипа Rhnull [501,647].
Определенную роль в происхождении Rhnull, как и других фенотипов делеций, играют браки между родственниками, способствующие накоплению гомозигот по депрессивным и молчащим аллелям. Как видно из приведенных выше данных, родители лиц Rhnullболее чем в половине случаев являлись кровными родственниками.
Молекулярно-генетические исследования, проведенные Cherif-Zahar и соавт. [210] и Huang и соавт. [357], выявили делецию гена RHD и 2 мутации, обусловливающие Rhnull аморфного типа. Выявлена делеция 2 нуклеотидов в кодо-нах 322 и 323 экзона 7 гена RHCE с нуклеотидной заменой ТСА Щ С. Такой ген производит укороченный протеин - 10 трансмембранных доменов из 398 аминокислот вместо 12 доменов из 417 аминокислот. Авторы полагают, что указанные изменения мешают взаимодействию Rh-протеина с Rh-ассоциированным гликопротеином (RhAG), в результате чего полноценный Rh-комплекс на поверхности мембраны эритроцита не образуется.
В одном случае Rhnull Huang и соавт. [358] нашли 2 перемещения G —> А в экзоне 6 гена RHAG (ассоциированного гликопротеина), которые приводили к замене Val 270 —| Не и Gly 280 й Arg. Другой случай Rhnull характеризовался трансверсией G —» Т в экзоне 9, которая вызывала замещение Gly 380 —> Val в трансмембранном сегменте 12.


{kind=link}