Поиск по сайту
- Вы здесь:
-
Главная

-
Медицинские автомобили
-
book
-
Группы крови человека
- Аллоиммунизация как глобальный популяционный процесс
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
Аллоиммунизация как глобальный популяционный процесс
Слабые варианты антигена В встречаются реже, чем А, преимущественно в монголоидных популяциях (Bhatia и соавт. [78], Lin-Chu и соавт. [144], Marsh и соавт. [150], Simmons и соавт. [196], Yamamoto [238], Yokoyama и соавт. [240], Yoshida и соавт. [246], Yu и соавт. [248]). Слабые варианты В труднее поддаются систематизации. Salmon (1976) предложил подразделять варианты В на В., В и Вт. Другие специалисты считают такую номенклатуру неудачной, поскольку она формально повторяет классификацию подгрупп А (А3, Ах, А ), но на практике ей не соответствует. Другой подход, основанный на подсчете процента агглютинированных и неагглютинированных эритроцитов, предложили Lopez и соавт. в 1973 г. Авторы подразделили варианты В на В60, В20 и В.
Экспрессия антигена В выше у негроидов, чем у лиц белой расы.
Вз
Вариант В3 описали Wiener и Cioffi (цит. по Daniels [87]). Эритроциты пробанда показывали смешанный характер агглютинации с антителами анти-В и анти-А,В. В слюне присутствовало вещество В. Фенотип В3 зафиксирован у французов с частотой 1 на 10 000, у китайцев - 1 на 900 среди лиц группы В и 1 на 1800 среди лиц группы АВ, в последнем случае фенотип соответствовал А,В3 (Lin-Chu и соавт. [144]). Посемейные исследования подтвердили наследственную передачу признака В3 как продукта редкого аллеля АВО-локуса. У лиц подгруппы В3 В-трансфераза присутствовала в сыворотке крови, на эритроцитах ее не выявили (Lin-Chu и соавт. [144]).
Для эритроцитов Вх характерны слабые реакции с антителами анти-В и анти-А,В. Сыворотка крови лиц Вх содержит слабые антитела анти-В, в секретах выделителей имеется небольшое количество вещества В. Посемейными исследованиями показана передача гена Вх по наследству как редкого аллеля АВО- локуса. В-трансфераза в сыворотках крови людей подгруппы Вх не выявлена.
Вт
Эритроциты этой подгруппы не агглютинируются антителами анти-В и анти-А,В. Антиген В выявляют методом адсорбции - элюции. В сыворотке крови лиц Вщ анти-В-антитела обычно отсутствуют. Содержание В-трансферазы в сыворотке крови сниженно, на эритроцитах - в следовом количестве (Gundolf и соавт. [114]).
в“
Эритроциты Ве1 не агглютинируются антителами анти-В и анти-А,В; вещество В в секретах отсутствует, сыворотки крови иногда содержат антитела анти-В. В-трансфераза не выявлена ни в сыворотке крови, ни на эритроцитах. Прослежена наследственная передача гена Bel.
Bw
Фенотип нескольких лиц, имевших эритроциты со слабой экспрессией антигена В, названный Bw, не удалось идентифицировать серологическими методами. Различия с другими вариантами В выявлены только молекулярногенетическими методами.
Аmos
Выявлены лица группы А, эритроциты которых проявляли мозаичный (mosaic - отсюда и обозначение) характер агглютинации со специфическими антителами. Эритроциты Ams, будучи смешанными с реагентами анти-А, отличались от А3 и Ах тем, что процент неагглютинированных клеток был постоянным и не зависел от активности анти-А-антител в тест-системе. Эритроциты, не вовлеченные в агглютинацию, не обладали способностью адсорбировать анти- А-антитела. Активность сывороточных А-трансфераз у лиц Amos снижена.
Возникновение варианта Amos связывают с соматическими мутациями в отдельных клонах гемопоэтических клеток (Marsh и соавт. [151]), в результате чего наряду с эритроцитами, содержащими антиген А, продуцируются эритроциты, лишенные этого антигена.
Ат
Фенотип А , описаный Gammerlgaard в 1942 г., сначала был обозначен им как Ах, но, поскольку найденный вариант отличался от А , вновь открытую подгруппу переименовали в Ат. Эритроциты Ат реагируют очень слабо и не со всеми образцами антител анти-А и анти-А,В. Присутствие антигена А в эритроцитах устанавливают с помощью адсорбции - элюции. У лиц А выделителей количество вещества А и Н в секретах приближается к норме. Сыворотки их крови, как правило, не содержат антител анти-Аг При посемейных исследованиях установлено, что ген Ат передается по наследству как редкий аллель АВО. Среди французов данный фенотип встречался с частотой 1 на 150 тыс. (Damborough и соавт. [88]), среди китайцев Тайваня - 1 на 400 тыс. (Daniels [87], Damborough и соавт. [88]). Несмотря на редкость фенотипа Ат, удалось идентифицировать 2 его разновидности с помощью детального анализа свойств сывороточных а-1,3 -ТУ-галактозаминилтрансфераз. Активность этих ферментов отличалась по кинетическим характеристикам при определенных значениях pH. В целом активность А-трансфераз у лиц подгруппы Аю ниже, чем у лиц Aj. Найдены лица группы АщВ. Посемейные исследования не позволили установить наследственную передачу Ат-фенотипа.
Ау
Подгруппа Ау напоминает Ат, однако имеет отличия:
-элюаты анти-А-антител, снятые с эритроцитов Ау, менее активны по сравнению с элюатами, снятыми с эритроцитов Ат;
- слюна лиц Ау выделителей содержит существенно меньше вещества А по сравнению с лицами Ат;
h -- сыворотка крови людей Ау содержит следовое количество А-трансферазы, а у индивидов Ат этот фермент отчетливо выявляют (Drozda и соавт. [92], Issitt, Anstee [122], Reid, Lomas-Francis [186]).
Полагают, что фенотип Ау не передается по наследству, а возникает в результате мутации зародышевых клеточных линий в пределах одной и той же семьи.
Aei
Подгруппа Ае1 (elution) названа так по методу, с помощью которого удается обнаружить присутствие антигена А на эритроцитах. Антитела анти-А и анти- А,В Не агглютигнируют эритроциты Ае1, однако адсорбируются на их поверхности, что подтверждают методом адсорбции - элзоции. В сыворотках крови лиц Ае1 А-трансфераза не выявляется, содержание Н-трансферазы снижено.
Фенотип Ае1 встречается редко: 1 на 100 тыс обследованных, преимущественно среди монголоидов (Solomon и соавт. [197], Sturgeon и соавт. [208]). Посемейными наследованиями показана передача гена Ael как редкого аллеля локуса АВО.
Эти варианты антигена А отличаются крайне слабой экспрессией (Mohn и соавт. [157], Nevanlinna и соавт. [168], Sturgeon и соавт. [208]), поэтому и обозначены как «end».
Антиген А . практически не реагирует с антителами анти-А,В лиц группы О, не взаимодействует с антителами анти-А лиц группы В. Некоторое количество группового вещества А выявляют в слюне.
Подгруппа найдена всего дважды среди 150 тыс. французских доноров.
Другой, несколько отличающийся от Aend, вариант был описан у финнов, в связи с чем и получил обозначение Afinn. Его находили с частотой 1 на 1000— 6000 обследованных в различных районах Финляндии (Mohn и соавт. [157]). Агглютинация эритроцитов Afinn с антителами анти-А и анти-А,В различима лишь при микроскопировании. В одном поле зрения насчитывают от 2 до 10 агглютинатов, каждый из которых состоит из 4-6 эритроцитов (Mohn и соавт. [157]). Слюна выделителей содержит вещество Н, вещество А отсутствует. Экстраагглютинины анти-Aj выявлены в сыворотках крови всех лиц Afinn. Этим подгруппа Afinn отличается от других вариантов Aend. Issitt и Anstee [122] считают выделение подгруппы Afinn в качестве самостоятльной недостаточно обоснованным.
Еще один вариант A d, получивший обозначение Аbantu, выявлен в Южной Африке у негроидов банту с частотой 4-8 % (Nevanlinna и соавт. [168], Sturgeon и соавт. [208]). Эритроциты Abantu реагируют с антителами анти-А несколько интенсивнее, чем А .. Сыворотки крови некоторых лиц Abantu содержали экстраагглютинины анти-Аг Посемейные исследования подтвердили, что аллель Аbantu передается по наследству. Высказано предположение, что в негроидной популяции этот ген появился сравнительно недавно вследствие миграции населения Африки в южном направлении. Он практически отсутствует среди негроидов Западной Африки и их потомков, проживающих в настоящее время в Америке.
Ainc
Название антигена «1ае» происходит от аббревиатуры слов «лектин», «адсорбция», «элюция».
Вариант А1ае описан Schuh, Vyas, Fudenberg в 1972 г. у 9 членов одной французской семьи. Присутствие антигена А в эритроцитах А1ае было установлено с помощью адсорбции и элюции лекгина анти-Aj из Dolichos biflorus. В слюне субстанция А отсутствовала. Сыворотки крови лиц А1ае реагировали с эритроцитами А,, \ и В.
Считается, что указанный фенотип это результат действия редкого гена в локусе АВО.
Вариант А1ае является, по-видимому, первым промежуточным субстратом между антигенами О и А. Об этом свидетельствует тот факт, что эритроциты OCad+, так же как А, , способны, хотя и в меньшей степени, адсорбировать лектин Dolichos biflorus.
Араc
Обозначение «рае» также представляет собой аббревиатуру: р - лектин из белковых желез улитки Helix pomatia, с помощью которого антиген А выявляют на эритроцитах, ае - метод детекции: адсорбция, элюция.
Подгруппа Арае была обнаружена в 1987 г. Stemps и соавт. сразу в 3 семьях. Предполагают, что фенотип А является вариантом подгруппы Ах и АЬе.
По мнению классиков клинической трансфузиологии (Mollison и соавт. [159]), гемолитические посттрансфузионные реакции в таких случаях могут иметь место, но их выраженность невелика по сравнению с другими иногрупп- ными трансфузиями, и они легко купируются. Эритроциты А,, несущие небольшое количество А-эпитопов, менее чувствительны к повреждающему действию специфических антител. Клинически значимый гемолиз in vivo развивается лишь в тех случаях, когда анти-А-антитела в сыворотке крови реципиента имеют высокую активность и титр (1 : 1000 и выше), что встречается редко.
С начала 2000-х годов некоторые специалисты службы крови поднимают вопрос о необходимости учета подгрупп А при переливании эритроцитсодержащих компонентов крови. Распространяются реагенты для идентификации подгрупп и рекомендации переливать лицам А2 только эритроциты А2, а лицам А2В - эритроциты А2В. Указанные рекомендации не имеют научного обоснования.
За последние 50 лет в Российской Федерации выполнено не менее 30 млн трансфузий эритроцитов группы А реципиентам группы А, из которых 12 % (согласно статистике) имели подгруппу А2. Ни одного случая посттрансфузион- ного осложнения, обусловленного различием донора и реципиента по подгруппе А, не зафиксировано.
Эритроциты А(П) с пониженными агглютйнационными свойствами впервые описали Dungem и Hirszfeld в 1911 г. [94]. Landsteiner и Levine [133] подразделили фенотип А на подгруппы.
Около 88 % лиц группы А относятся к подгруппе А1 12 % - к подгруппе А^. Среди лиц AB(IV) 80 % Afi, 20 % А2В. Ранее полагали, что лица А1 имеют антигены А и Ар а лица А2 - только антиген А. Клетки и А2 несут различное количество антигенного вещества, имеющего в то же время и некоторые качественные отличия. Так, сыворотки анти-А лиц, имеющих группу крови В(111), содержат 2 фракции анти-А-антител: одна фракция (анти-А) реагирует с эритроцитами Aj и А2, другая (анти-Aj) - только с эритроцитами кх (Wiener и соавт. [225, 227]). Адсорбция сывороток анти-А эритроцитами А2 позволяет получить реактив, который не реагирует с эритроцитами А2, но сохраняет высокую активность по отношению к эритроцитам и может быть использован для дифференцировки образцов эритроцитов А на Аги А2(Р.М. Уринсон [61]).
Для определения подгрупп А используют экстраагглютинины и а2, специально отобранные моноклональные антитела анти-А, а также лектины Dolichos biflorus и Ulex europaeus. Первый из них реагирует с эритроцитами А1 второй - с эритроцитами А2.
У лиц Aj с частотой примерно 1 на 10 ООО встречаются экстраагглютинины о^. Лица А2 и А2В содержат экстраагглютинины cL с частотой 1 на 50 и 1 на 5 соответственно.
Гены А1 и А2 передаются по наследству. Лица А1 А2 имеют фенотип Аг
Возникновение 2 или более популяций эритроцитов, несущих различное количество А-эпитопов, обусловлено А-трансферазами, модифицировавши вирусами или сформировавшимися под влиянием атипичных 50-аллелей.
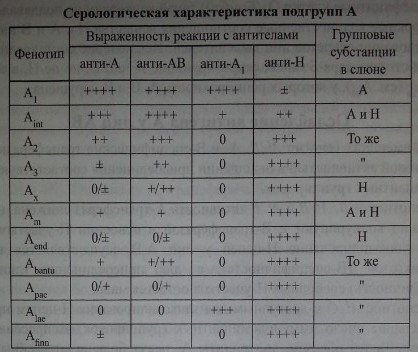
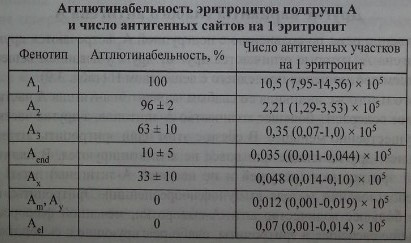
Агглютинационные свойства эритроцитов различных подгрупп А, а также число А-эпитопов весьма вариабельны.
Варианты антигена А получили индивидуальные названия, однако это не означает, что каждый их них представляет независимую генетически детерминированную подгруппу. Они скорее являются исключением из правил нормального формирования группового вещества.
Как уже отмечалось, большинство антител анти-А лиц группы В не агглютинирует эритроциты Ах. Сыворотки крови лиц В, иммунизированных групповым веществом А, напротив, обладают такой способностью. Моноклональные реагенты анти-А выявляют подгруппу Ах, однако в некоторых случаях эти тест- системы взаимодействуют также и с антигеном В.
Эритроциты Ах очень редки - 1 : 40 000-77 ООО (Reid, Lomas-Francis [186]).
Специфическая А-трансфераза обычно отсутствует в сыворотках крови и на эритроцитах лиц подгруппы А , активность Н-трансферазы снижена.
Результаты посемейных исследований противоречивы. В некоторых семьях удавалось проследить наследование подгруппы Ах как фенотипического проявления редкого аллеля А . В других семьях пробанды Ах имели родителей О (Beckers и соавт. [73], Valdes и соавт. [214], Van Loghem и соавт. [215]). Ducos и соавт. [93] наблюдали детей Ах в семьях, где один из родителей имел группу А2В.
Дифференцировать подгруппы А2/В и Ах/В можно по отсутствию в сыворотке крови лиц Ах/В A-специфической трансферазы.
Как отмечают Issitt и Anstee [122], некоторые производители моноклональных антител предъявляют повышенные требования к своим реактивам и используют для оценки их качества эритроциты А .
Вещества, вызывающие агглютинацию А-, В- или Н-положительных эритроцитов человека, были найдены во многих растениях, водорослях, грибах, а также в тканях некоторых червей, улиток, икре лососевых рыб, сыворотке крови угрей. Они были названы лектинами (табл. 3.6). Установлено, что они взаимодействуют с терминальными моносахаридными группировками. Лектины из Dolichos biflorus и Ulex europaeus используют для дифференцировки подгрупп А в исследовательских целях (П.Н. Косяков [35], Прокоп, Геллер [56], Daniels [87], Issitt, Anstee [122], Sringarm и соавт. [201]), в рутинной практике определения групп крови АВО их не применяют.
Таблица 3.6
Лектины с анти-А-, анти-В-, анти-AB- и анти-Н-активностью
|
Источник получения |
Специфичность |
|
|
Dolichos biflorus |
Семена |
Анти-А |
|
Phaseolus limensis |
To же |
Анти-А |
|
Phaseolus lunatus |
II |
То же |
|
Helix pomatia |
Печень |
II |
|
Helix hortensis |
To же |
II |
|
Fomes fometarius |
Мицелий |
АнпьВ |
|
Ptilota plumosa |
Органеллы |
То же |
|
Salmo salar |
Икра |
)1 |
|
Sophora japonica |
Семена |
Анти-АВ |
|
Phlomis Jructosa |
То же |
То же |
|
Bandeiraea simplicifolia |
II |
II |
|
Ulex europaeus |
II |
Анти-Н |
|
Lotus tetragonolous |
11 |
Анти-Н (HI) |
|
Anguilla anguilla |
Анти-Н (HI) |
|
|
Cystisus sessifolius |
Семена |
Анти-Н |
|
Laburnum alpinum |
То же |
То же |
Антиген С
Существование антигена С в системе АВО можно считать доказанным. Он содержится в эритроцитах А(П), В(Ш) и AB(IV). В эритроцитах 0(1) антиген С отсутствует.
Основанием для такого утверждения служат эксперименты Wiener и соавт. [213, 224, 228-230], показавшие, что сыворотки Оар(1), помимо а- и Р-агтлютининов, реагирующих с эритроцитами А и В, содержат еще одну разновидность естественных групповых антител - С-агтлютинины (по Винеру анти-С- антитела), которые реагируют с эритроцитами А и В перекрестно
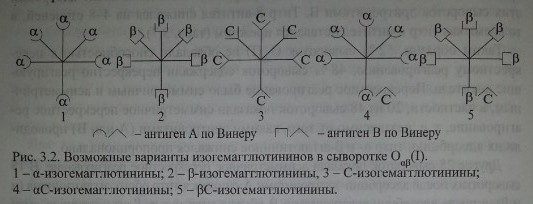
На наш взгляд, перекрестное реагирование обусловлено не ар-агглютининами, как полагала Dodd и соавт., а антителами аС, PC, что в большей мере согласуется с концепцией Винера, а также полученными нами данными. На рис. 3.2 представлена предполагаемая модель этих антител.
Представленная структурная модель изогемагглютининов позволяет объяснить характер перекрестного реагирования сывороток Оар(1). Сыворотки, не вызывающие перекрестных реакций, содержат только а- и (3-антитела (рис. 3.3, п. 1). Сыворотки, вызывающие симметричные перекрестные реакции, содержат, помимо а- и р-антител, антитела аС и PC или анти-С-антитела (рис. 3.3, п. 4). Сыворотки, вызывающие асимметричные перекрестные реакции, содержат а, р и одну из разновидностей антител - аС или PC (рис. 3.3, п. 2, 3).
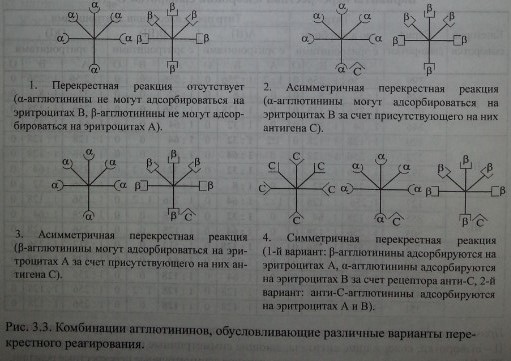
Комбинации агглютининов, обусловливающие различные варианты перекрестного реагирования.
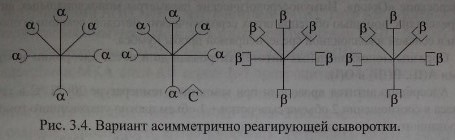
Вариант асимметрично реагирующей сыворотки.
Согласно схеме сыворотка содержит 50 % а-агглютининов, 75 % - р. Валентность анти-С реагирует как а и р. При адсорбции такой сыворотки эритроцитами А а-агглютинины будут удалены полностью. Иными словами, из сыворотки будет удалено 50 % антител. При адсорбции сыворотки эритроцитами В будет удалено 75 % антител (50 % Р+25 % а) за счет рецептора анти-С. В случае, если рецептор анти-С связан не с а-, а с р-агглютининами, асимметричное реагирование проявят эритроциты А, унося на своей поверхности 25 % Р-агглютининов.
Как далее нами показано, перекрестно реагирующие антитела так же, как иммунные антитела АВО и Rh, относятся к классу IgG, устойчивы к унитиолу, лучше реагируют с отмытыми, чем неотмытыми эритроцитами, в отличие от иммунных (термо- сгабильных) антител являются термолабильными (утрачивают активность после прогревания сыворотки при 70 °С в течение 10 мин). В противоположность классическим иммунным антителам IgG, которые адсорбируются на эритроцитах, но не вызывают их агглютинации, перекрестно реагирующие антитела непосредственно агглютинируют эритроциты в солевой среде. В низкой концентрации они утрачивают способность агглютинировать эритроциты, однако сенсибилизируют их подобно неполным антителам и хорошо выявляются в непрямой антиглобулиновой пробе Кумбса.
Сыворотки лиц 0^(1), очевидно, содержат несколько типов групповых антител: изогемагглютинины аир IgM, аир IgG, аС и рС IgG* анти-С IgG, иммунные аир IgM, иммунные а и Р IgG, что закономерно обусловливает перекрестные реакции с различными групповыми антигенами.
Важным аргументом, подтверждающим концепцию Винера о существовании антигена С, явились эксперименты по получению специфических антисьгворотк. В частности, при иммунизации мышей эритроцитами А(П) и В(Ш) нами [21] были получены моноклональные антитела со специфичностью анти-АВ (МКА-АВ). Материал и методы. Трехмесячным самкам мышей BALB/c вводили по 0,1 мл отмытых 0,9% NaCl эритроцитов А(П) или В(Ш) внутрибрюшинно дважды с интервалом в 7 дней. Третью инъекцию производили внутривенно за 5 дней до слияния спле- ноцитов с клетками миеломы NS-1. Слияние осуществляли по стандартной методике с использованием 45% полиэтиленгликоля-1500 «ЬоЬа» и 5% диметилсульфоксида.
Эффективность гибридизации составляла 10-5. Клонировали гибридные клетки в среде HAT без фидера. Антителопродуцирующие клоны отбирали с помощью реакции аг
в 96-луночных планшетах с отмытыми эритроцитами 0(1), А(П) и В(Щ), Класс моноклональных антител устанавливали по флюоресценции сенсибилизи-IgG, нейтрализовались группоспецифическими субстанциями, существенно отличает их от классических иммунных антител и позволяет выделить их в особую группу - иммунные IgG-антитела термолабильные легко нейтрализуемые. К этой же группе могут быть отнесены перекрестно реагирующие антитела лиц 0(1), имеющие идентичную иммуносерологическую характеристику - IgG термолабильные. Их происхождение (иммунное или естественное) остается неясным. Не исключено, что они так же, как МКА анти-АВ, являются иммунными.
Обработка эритроцитов В(Ш) протеолитическими ферментами, специфически разрушающими антиген В, лишала эритроциты групповых свойств. После такой обработки эритроциты В(Ш) в отличие от обработанных глутаровым альдегидом приобретали свойства эритроцитов 0(1) группы и не агглютинировались сыворотками анти-В. Одновременно исчезала их способность взаимодействовать с перекрестно реагирующими антителами МКА анти-АВ, а также перекрестно реагирующими антителами лиц 0(1).
По характеру реагирования полученные МКА анти-АВ вряд ли можно отнести к анти-А и анти-В. По-видимому, это одно, маскирующееся под анти-А и анти-В антитело, реагирующее с общей для эритроцитов А(П) и В(Ш) антигенной детерминантой. По своим свойствам эти антитела близки к перекрестно реагирующим антителам сывороток лиц 0(1), описанным выше. Сам факт получения антител анти-АВ в ответ на иммунизацию эритроцитами как А, так и В служит веским аргументом в пользу того, что оба антигена наряду с существенными серологическими различиями имеют идентичный эпитоп С, в равной мере являющийся сильным иммуногеном.
Таким образом, имеются все основания полагать, что групповая система АВО представлена не двумя агглютиногенами - А, В и двумя агглютининами - а, Р, а тремя агглютиногенами - А, В, С и тремя агглютининами - а, р и С.
Антиген С выявляют с помощью перекрестно реагирующих сывороток лиц Оар(1) [184, 229]. Именно фракция перекрестно реагирующих агглютининов представляет собой антитела анти-С. Последние можно удалить из сыворотки указанных лиц адсорбцией как А, так и В эритроцитами. При рутинном исследовании антиген С маскируется присутствием антигена А и/или В. Антитела анти-С в обычных методах исследования проявляют себя подобно несепари- руемой комбинации а- и Р-агглютининов. У лиц, имеющих группу крови А и В, агглютинины анти-С отсутствуют.
Не исключено, что антитела анти-С могут встречаться в чистом виде. Однако отличить их от анти-А и анти-В обычными методами не представляется возможным. У плодов 0(1) от матерей 0(1) находят антитела со свойствами анти-С, у плодов А(Н) и В(Ш) от матерей А(Н) и В(Ш) таких антител не находят.
Owen (цит. по Wiener [229]) получил чистую фракцию анти-С-агглютининов адсорбцией сывороток 0^(1) эритроцитами опоссума и кролика, которые, содержат парциальный антиген В и, по-видимому, не содержат антигена С. После такой обработки оставшиеся в сыворотке антитела анти-С реагировали с эритроцитами А и В, и их можно было удалить эритроцитами А или В в ад- <юрбционной пробе. Однако для окончательного доказательства того, что С-, о- и Р-агглютинины являются самостоятельными независимыми друг от друга антителами, не достает эксперимента, в котором антигены А и В были бы разрушены ферментативно (не реагировали бы с сыворотками анти-А и анти-В), но при этом сохранили бы антиген С (продолжали бы агглютинироваться анти-С- сывороткой или элюатом аР). Только в том случае, если эритроциты не агглютинируются а и р, но продолжают агглютинироваться анти-С-антителами, можно не только с достаточной степенью уверенности констатировать существование антигена С, но и с помощью такого реактива изучить закономерности наследования этого антигена, возможные его разновидности и комбинации. В наших экспериментах ферментативное разрушение В-антигена приводило одновременно к утрате их способности реагировать с МКА анти-АВ. Иными слова ми разрушение В-антигена влекло за собой разрушение антигена С. Однако, не смотря на отрицательный результат этого эксперимента, полученные нами данные в целом со всей очевидностью свидетельствуют о наличии третьего антигена в системе АВО, существование которого блестяще предсказал Wiener. глютинации
Изогемагглютинины анти-А и анти-В отсутствуют в крайне редких случаях: у больных врожденной гипо- и агаммаглобулинемией, а также при Х-ассоциированном синдроме Вискотта - Олдриджа. Такие лица не способны синтезировать антитела к полисахаридным, но не белковым антигенам.
Изогемагглютинины отсутствуют при истинном химеризме, наблюдаемом у ди- зиготных близнецов. В период внутриутробного развития между близнецами может происходить обмен кроветворными клетками, в результате чего после рождения у каждого из близнецов в кровотоке присутствуют 2 популяции эритроцитов с различными фенотипами. Описаны химеры, когда 90 % циркулирующих эритроцитов имели группу О, а остальные 10 % - группу А. У таких индивидов в плазме крови обнаруживали только анти-В-антитела, анти-А отсутствовали (см. Кровяные химеры).
В очень редких случаях изогемагглютинины отсутствовали у здоровых лиц без признаков гипогаммаглобулинемии или какой-либо другой патологии. По данным Dobson и Ikin, изогемагглютинины отсутствовали у 0,01 % здоровых лиц. Авторы связывали это с соматическими мутациями, приводящими к инактивации клонов В-клеток, запрограммированных на синтез соответствующих антител. Интересно, что родители, братья и сестры пробандов содержали изогемагглютинины, характерные для соответствующей группы крови. Возможно, синтез АВО-антител контролируется широко встречающимся доминантным геном и его редко встречающимся рецессивным молчащим аллелем. Лица без изогемагглютининов являются гомозиготными по указанному рецессивному аллелю.
Онтогенез АВО-антигенов
Подобно антителам, антигены системы АВО в момент рождения развиты не полностью, хотя у эмбрионов их обнаруживают уже на 5-й неделе внутриутробного развития. Эритроциты пуповинной крови реагируют с антителами анти-А и анти-В слабее по сравнению с клетками взрослых лиц (Witebsky и Engasser [97, 233], Romano и соавт. [187]). Эритроциты некоторых новорожденных группы А по своей агглютинабельности могут приближаться к эритроцитам А2. У 6-18-месячного ребенка агглютинабельности эритроцитов повышается, серологические характеристики соответствуют группе А,. Эритроциты новорожденных А, лучше реагируют с лектином Dolichos biflorus, чем с антителами анти-А, аллогенного происхождения. Эритроциты детей А2 более интенсивно взаимодействуют с антителами анти-А,В, чем с антителами анти-А, и тем самым показывают сходство с подгруппой Ах. Антиген Н на эритроцитах новорожденных выражен почти так же, как и у взрослых.
Эритроциты новорожденных А несут существенно меньшее количество А-антигенных участков (табл. 3.5) по сравнению с клетками взрослых лиц А, (Economidou и соавт. [95]). В то же время более высокая активность ^-трансферазы зарегистрирована в сыворотках пуповинной крови, чем в сыворотках крови взрослых.
Слабые реакции эритроцитов новорожденных с антителами анти-А и анти-В можно объяснить тем, что указанные антитела связываются с эритроцитами не двумя, а одним Fc-учаетком. На эритроцитах детей цепи, несущие антигены А и В, недостаточно разветвлены. По мере разветвления меняется пространственная конфигурация эпитопов, их количество увеличивается и они становятся доступнее для активных участков антител. Повышение агглютинабельных свойств эритроцитов происходит постепенно в течение первых 6-18 мес. жизни. Концентрация (31 —»6-глюкозаминилтрансферазы, участвующей в формировании разветвления, у новорожденных низкая. Содержание этого фермента достигает уровня его у взрослых в течение первых 6-18 мес. жизни.
Таблица 3.5
Плотность распределения А-антигена на эритроцитах
|
Группа крови |
Количество антигенных участков, тыс. на 1 эритроците |
|
|
новорожденных |
взрослых |
|
|
Ai . |
250-370 |
810-1170 |
|
А2 |
140 |
240-290 |
|
А,В |
220 |
460-850 |
|
а2в |
|
120 |
|
В |
|
610-830 |


{kind=link}