Поиск по сайту
- Вы здесь:
-
Главная

-
Медицинские автомобили
-
book
-
Группы крови человека
- Система Cartwright и система Xg
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
Система Cartwright и система Xg
Антитела aHTH-Xga выявлены Mann и соавт. [64] в сыворотке крови мужчины, которому многократно переливали кровь. Позднее найдены другие образцы антител указанной специфичности (Azar и соавт. [4], Cook и соавт. [17], Devenish и соавт. [27], Herron, Smith [53], Мак и соавт. [63], Metaxas, Metaxas-Buhler [66], Nakajima и соавт. [67], Race, Sanger [73], Sausais и соавт. [82], Yokoyama и соавт. [99]).
AHTH-Xga-amir^a встречаются редко: при исследовании 325 сывороток в Гонконге антитела anra-Xg3 были идентифицированы в 4 случаях, один образец указанных антител приходился на 60 108 обследованных доноров (Azar и соавт. [4]).
Относительно низкая частота встречаемости антигена Xga среди мужчин дала основание полагать, что они в большей мере, чем женщины, подвержены аллоиммунизации антигеном Xga. Действительно, из 14 сывороток анти-Xga, найденных первыми, 12 образцов были получены от мужчин (Race, Sanger [73]), из 13 образцов анти-Xg3, выявленных у японцев, 11 принадлежали здоровым лицам - донорам мужчинам (Azar и соавт. [4]).
AHTH-Xga-aHTm^a чаще встречаются у монголоидов, что, по-видимому, обусловлено еще более низкой распространенностью антигена Xga среди представителей этой расы. Остается, однако, без объяснения тот факт, что антитела анти-Xg3 встречаются у мужчин значительно чаще, чем можно ожидать, исходя из частоты встречаемости этого антигена в популяции.
Антитела анти-Xg3 присутствуют в сыворотках обычно в чистом виде, без примеси других антител. Они, как правило, естественного происхождения: их обнаруживают у здоровых лиц, не имевших беременностей и гемотрансфузии. В большинстве случаев они относятся к классу IgG, и лишь отдельные образцы проявляют агглютинирующую активность в солевой среде. Адекватный метод выявления антиО(£а-антител - непрямая антиглобулиновая проба. Многие образцы указанных антител обладают способностью связывать комплемент. Посттрансфузионных осложнений и ГБН они не вызывают (Issitt, Anstee [56]). Описан больной, имевший aHTH-Xga-aHTm^a, которому были перелиты шесть доз эритроцитов Xg(a+) без каких-либо реакций (Cook и соавт. и соавт. [17]). Тесты с эритроцитами, меченными Сг51, подтвердили заключение о том, что aHTH-Xga-aHTm^a не влияют на продолжительность циркуляции перелитых эритроцитов (Sausais и соавт. [82]).
У одной женщины во время беременности выявлены аутоиммунные анти-Х§а-антитела (Yokoyama и соавт. [99,100]).
Ellis и соавт. [31] получили мышиные моноклональные aHTH-Xga-peareHTbi. Мышей предварительно иммунизировали субстратом, содержащим фрагменты N-терминального участка Xg-гликопептида.
В 1979 г. Levy и соавт. [60] путем иммунизации мышей лейкемическими Т-клетками человека получили моноклональные антитела анти-С099, которые, как показали Goodfellow и Tippett [48], реагировали с антигеном Xg.
Антиген CD99, открываемый этими антителам, присутствовал практически во всех тканях человека (Goodfellow [41]), однако в отличие от других клеток эритроциты экспрессировали его в разных количествах. С помощью радиоиммунного антиглобулинового метода и проточной цитофлюориметрии выделены два типа людей: с высокой (high) и низкой (low) экспрессией антигена CD99 на эритроцитах (Goodfellow, Tippett [48], Foucher и соавт. [36]).
Установлено, что высокая экспрессия CD99 характерна для эритроцитов Xg(a+). У женщин Xg(a-) экспрессия CD99 низкая. У 68 % мужчин Xg(a-) выявлена высокая экспрессия CD99, в 32 % - низкая.
На основании полученных данных Goodfellow и соавт. [45, 48], Tippett и соавт. [92] высказали предположение о существовании на Y-хромосоме локуса 7G, который контролирует уровень экспрессии антигена CD99. По мнению авторов, ген YG представлен двумя аллелями: Yga и Yg. Лица с высокой экспрессией CD99 имеют ген Xga и Yga, в то время как у людей со слабовыраженным антигеном CD99 присутствуют аллели Xga и Yg. Далее указанные авторы усовершенствовали предложенную модель, предположив существование гена XGR, регулирующего одновременно экспрессию антигенов CD99 и Xga и присутствующего как на Y-, так и на Х-хромосоме.
Uchikawa и соавт. [95] нашли у 2 японских доноров антитела анти-СБ99, подобные описанным выше, но несколько отличающиеся от них. Эти антитела реагировали с антигеном Xga, получившим обозначение XG2.
Вещество CD99, так же как Xga, относится к сиалогликопротеинам. Равным образом CD99 разрушается папаином, проназой, трипсином и химотрипсином (Goodfellow [41], Latron и соавт. [59], Daniels, Tippett [21]). Антиген CD99 в основном устойчив к действию сиалидазы, однако некоторые образцы анти-С099-подобных антител не реагировали с эритроцитами, обработанными сиалидазой (Goodfellow [41], Latron и соавт. [59], Daniels, Tippett [21]). Иммуноблоттинг с эритроцитами, лимфоцитами, различными клеточными линиями человека, гибридными клетками человек - мышь показал, что антиген CD99 ассоциирован с гликопротеином, имеющим мол. массу 32 кДа (Petty, Tippett [71], Fouchet и соавт. [37], Latron и соавт. [59], Daniels, Tippett [21], Banting и соавт. [6], Anstee и соавт. [3]). Обработка эритроцитов сиалидазой приводила к снижению мол. массы CD99 (Petty, Tippett [71], Fouchet и соавт. [37], Latron и соавт. [59], Anstee и соавт. [3]). Иммуноблоттинг преципитатов, полученных с помощью антител aHTH-Xga и анти-С099, показал, что эти антигены расположены на разных структурах (Fouchet и соавт. [37]). Частично очищенный гликопротеин CD99 ингибировал активность антител anra-CD99 и анти-Xg8 (Banting и соавт. [6]).
Petty и Tippett [71] установили, что аллогенные анти-Х§а-антитела преципитировали гликопротеины Xga и CD99. Авторы пришли к заключению, что эти высокогомологичные структуры связаны между собой и представлены в эритроцитарной мембране в виде гетеродимера. При использовании моноклональных aHTH-Xga-aHTm^ эти результаты воспроизвести не удалось (Fouchet и соавт. [37]).
Клонирование гена МС2 показало, что антиген CD99 локализован на протеине, состоящем из 186 аминокислот. Протеин включает N-терминальный сигнальный пептид из 20 или 21 аминокислоты (расщепляется после встраивания в мембрану эритроцита). Экстрацеллюлярная часть пептида включает 100 аминокислот, имеет гидрофобный трансмембранный домен и цитоплазматический фрагмент из 36 аминокислот (Goodfellow и соавт. [47], Banting и соавт. [5]). Различий между молекулами CD99, кодируемыми Х- и Y-хромосомами, не выявлено (Banting и соавт. [6]).
Подсчитано, что один лимфоцит несет 27 тыс. эпитопов CD99. Для тромбоцитов этот показатель составил 4 тыс. На эритроцитах число антигенных участков, связывающих анти-С099-антитела, существенно меньше: 1 тыс. для вариантов с высокой экспрессией CD99 и только 100 для вариантов с низкой экспрессией (Foucher и соавт. [36]).
Herron и Smith [53] посредством иммунопреципитации субстрата антителами анти-Xg3 установили, что антигенные эпитопы Xga эритроцитарной мембраны относятся к сиалогликопротеинам. Установлена мол. масса вещества (22,5-28 кДа). Обработка эритроцитов сиалидазой приводила к уменьшению мол. массы. Гликопротеин, определяющий специфичность Xg, тесно связан с белком CD99 (Petty, Tippett [71 ], Fouchet и соавт.
Кодируемый геном XG протеин состоит из 180 аминокислот. Экстрацеллюлярный N-терминальный домен включает 142 аминокислоты, содержит 16 участков О-гликозилирования, но не имеет участков N-гликозилирования. Остальная часть Xg-протеина представлена трансмембранным и црггоплазматическим С-терминальным доменами (рис. 14.1) (Ellis и соавт. [32]).
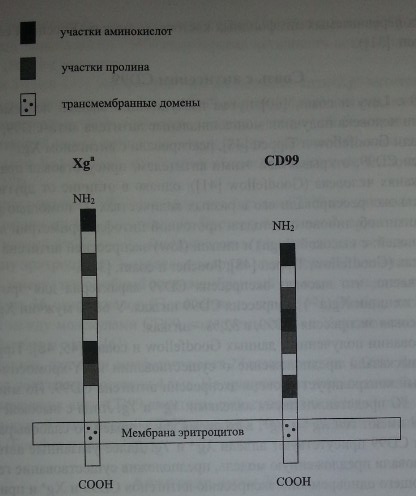
Строение гликопротеинов Xga и CD99
Строение гликопротеинов Xga и CD99
Транскрипты гена XG, помимо эритроидных клеток и фибробластов, выявлены в скелетных мышцах, сердце, плаценте, клетках предстательной и щитовидной желез, позвоночнике и трахее (Fouchet и соавт. [37]), Некоторое количество транскриптов XG обнаружено в легких, почках, яичке, а также в тканях плода и некоторых перевиваемых лимфоидных клеточных линиях.
Обследование 6784 жителей стран Северной Европы показало (табл. 14.2), что среди женщин частота фенотипа Xg(a+) составляет 89 %, в то время как среди мужчин - 66 % (Sanger и соавт. [80], Daniels [19], Haldane [51]).
Распределение антигена Xg у мужчин и женщин
|
Показатель |
Частота, % |
||
|
у мужчин |
у женщин |
||
|
Фенотип |
Xg(a+) |
65,9 |
88,7 |
|
Xg(a-) |
34,1 |
11,3 |
|
|
Генотип |
Xga/Xga |
43,4 |
|
|
XgVXg |
65,9 |
45,0 |
|
|
Xg/Xg |
34,1 |
11,6 |
|
Антиген Xga обнаружен у представителей всех исследованных популяций. Наибольшая частота гена Xgaконстатирована у аборигенов Новой Гвинеи и Австралии, жителей о. Сардинии и бразильцев ген Xga не вполне развит к моменту рождения. Его экспрессия на эритроцитах новорожденных ниже, чем на эритроцитах взрослых (Мауг [65], Toivanen, Hirvonen [94]). Вещество Xga формируется в относительно поздние сроки внутриутробного развития. Из 54 плодов от 6 до 20 недель развития только 19 были Xg(a+), что существенно ниже частоты встречаемости этого антигена в популяции (Toivanen, Hirvonen [93]). Антиген Xg(a+) появляется у плодов начиная с 12 недель развития. У 5-10 % мальчиков Xg(a+) при рождении указанный антиген не удается выявить серологическими методами (Szabo и соавт. [90]). По мере взросления организма выраженность его на эритроцитах возрастает (Сатрапа и соавт. [15]).
В процессе эритропоэза in vitro антиген Xga появлялся на клетках после гликофорина А и протеина полосы 3, но раньше Rh-протеина (Daniels, Green [20]).
Антиген Xga не является сильным иммуногеном. На эритроцитах гетерозиготных мужчин и гомозиготных женщин он выражен одинаково. У женщин, гетерозиготных по гену Xga, экспрессия антигена может быть снижена. До 10 % гетерозиготных женщин имеют слабовыраженный антиген Xga (Race, Sanger [73]). Слабые варианты Xga среди мужчин редки.
Количество антигенных участков Xga на эритроците, по данным разных авторов, варьирует от 159 (Foucher и соавт. [36]) до 9000 (Szabo и соавт. [90]).
В 1974 г. Fellous и соавт. [34] обнаружили антиген Xga на фибробластах и гибридомных клетках человек - мышь. Антиген выявлялся одновременно с другими продуктами Х-ассоциированных локусов. Тем самым было еще раз подтверждено, что антиген Xga контролируется геном, расположенным на Х-хромосоме. При инактивации Х-хромосомы антиген Xga на клетках человека не выявлялся (Hsu и соавт. [54]).
Антиген Xga разрушается бромелином, фицином, папаином, проназой, трипсином и химотрипсином (Habibi и соавт. [49], Herron, Smith [53]), однако устойчив к воздействию сиалидазой.
Частота генов XG у разных народов
|
Популяция |
Количество обследованных |
xga1 |
Xg |
Источник |
|
Аборигены Новой Гвинеи (папуасы) |
263 |
0,85 |
0,15 |
[85] |
|
Австралийские аборигены |
352 £ |
0,79 |
,0,21 |
[85] |
|
Индейцы навахо |
308 |
0,77 |
0,23 |
[28] |
|
Жители о. Сардиния |
322 |
0,76 |
0,24 |
[89] |
|
Бразильцы белые |
1 078 |
0,74 |
0,26 |
[68] |
|
Бразильцы мулаты |
786 |
0,62 |
0,38 |
[68] |
|
Бразильцы негры |
827 |
0,57 |
0,43 |
[68] |
|
Евреи Израиля, сефарды |
201 |
0,68 |
0,32 |
[1] |
|
Японцы |
529 |
0,68 |
0,32 |
[67] |
|
Жители стран Северной Европы |
15 716 |
0,66 |
0,34 |
[16,27,52,65,66, 80] |
|
Индусы, Бомбей |
100 |
0,65 |
0,35 |
[10] |
|
Индусы, Сингапур |
|
0,57 |
0,43 |
[77] |
|
Китайцы, континентальный Китай |
171 |
0,60 |
0,40 |
[28] |
|
Китайцы, Тайвань |
178 |
0,53 |
0,47 |
[28] |
|
Китайцы, Гонконг |
1300 |
0,49 |
0,51 |
[63] |
|
Китайцы, Сингапур |
165 |
0,45 |
0,55 |
[11,77] |
|
Испанцы |
636 |
0,59 |
0,41 |
[96] |
|
Таицы |
181 |
0,57 |
0,43 |
[74] |
|
Греки |
638 |
0,55 |
0,45 |
[38] |
|
Негры, Нью-Йорк, жители Ямайки |
219 |
0,55 |
0,45 |
|
|
Малайцы, Сингапур |
72 |
0,54 |
0,46 |
[77] |
|
Тайваньцы (аборигены) |
164 |
0,38 |
0,62 |
[28] |
Хромосомы, определяющие пол, так же как и аутосомы, принимают участие в формировании групповых признаков крови человека. В 1962 г. были открыты группы крови, которые передаются по наследству с Х-хромосомой.
Mann и соавт. [64] выявили в сыворотке крови мужчины белой расы (мистера And) необычные антитела, которые в отличие от всех известных ранее существенно чаще реагировали с эритроцитами женщин, чем мужчин. Посемейные исследования подтвердили, что антиген, получивший обозначение Xga, находится под контролем гена Xga (XG1\ сцепленного с Х-хромосомой. Поскольку антиген, антитетичный антигену Xga, не найден, было сделано заключение, что систему Xg образуют два гена: продуктивный ген-Xga(XGl) и его молчащий аллель-Xg (XG0).
С группами крови Xg тесно связан антиген CD99, находящийся под контролем гена МС2, присутствующего как на Х-, так и на Y-хромосоме (Goodfellow, Tippett [48]). В связи с этим антигены Xgaи CD99 оказались весьма ценными маркерами при изучении сцепленности различных признаков с полом, X-Y-гибридизации и рекомбинации, инактивации Х-хромосомы, Х- и Y-анэуплоидии, патологии формирования пола, в частности мужского типа у лиц XX.
Антиген Xga
Наследование
Пол человека детерминирован двумя хромосомами: XX у женщин, XY у мужчин. Ген Xga (XG1) расположен на Х-хромосоме, на Y-хромосоме он отсутствует. Если на Х-хромосоме мужчины присутствует аллель Xga, то он имеет фенотип Xg(a+). Если на Х-хромосоме мужчины отсутствует аллель Xga, а имеется молчащий ген Xg9 то формируется фенотип Xg(a-). Женщины Xg(a+) могут быть гомозиготными (Xga/Xga) или гетерозиготными (Xga/Xg\ в то время как все мужчины Xg(a+) гетерозиготны (Xga/Xg). Соответственно антиген Xga чаще встречается у женщин.
Анализ распределения групп крови Xg в 2540 семьях с 5824 детьми в Северной Европе (Sanger и соавт. [80]), а также во многих других семьх, включая жителей Канады (Chown и соавт. [16]), Сардинии (Suniscalo и соавт. [89]), Израиля (Adam и соавт. [1]) и Японии (Nakajima и соавт. [67]), подтвердил, что антиген Xga передается по наследству как Х-сцепленный доминантный признак. Исключения из этого правила, т. е. передача гена Xga с Y-хромосомой или другим путем, очень редки.
Таблица 14 J
Варианты фактического наследования антигена Xg
|
Фенотипы и генотипы |
|||||||
|
родителей |
детей |
||||||
|
отца |
матери |
сына |
дочери |
||||
|
Xg(a+) |
Xga |
Xg(a+) |
Xga/Xga |
Xg(a+) |
Xga |
Xg(a+) |
Xga/Xga |
|
Xg(a+)' |
Xga |
Xg(a+) |
Xga/Xg |
Xg(a-)1 |
Xg |
Xg(a+) |
XgVXg |
|
Xg(a-)2 |
Xga |
Xg(a-) |
Xg/Xg |
Xg(a+) |
Xga |
Xg(a+)2 |
Xg/Xga |
|
Xg(a-)3 |
Xg |
Xg(a+) |
Xga/Xga |
Xg(a+) |
Xga |
Xg(a+) |
Xg/Xga |
|
|
|
|
|
Xg(a-)3 |
Xg |
Xg(a-)3 |
Xg/Xg |
|
Xg(a-) |
Xg |
Xg(a-) |
Xg/Xg |
Xg(a-) |
Xg |
Xg(a-) |
Xg/Xg |
1 Данные противоречат положению о доминантном типе наследования Xga,
2 Ген Xgf2 фенотипически себя не проявляет, но передается по наследству,
3 Ожидаемый антиген Xga отсутствует.
В некоторых семьях не наблюдали Х-сцепленного наследования (табл. 14.1). Обычно мужчины Xg(a+) наследуют ген Xg61 от матери. Однако в 16 семьях сыновья женщин Xg(a-) имели группу Xg(a+) (Sanger и соавт. [78, 80], Chown и соавт. [16], Tippett, Ellis [91], Race, Sanger [73]).
Race и Sanger [73] высказали предположение, что небольшая часть Х-хромосомы, включающая участок XG, может транслоцироваться на Y-хромосому. Далее реком-бинантная Y-хромосома передается по наследству сыновьям. Таким образом, антиген Xg в части случаев может передаваться с Y-хромосомой. Однако и такой механизм наследования не объясняет встречающиеся варианты. В одной семье у мужчины Xg(a+) мать была Xg(a-) и его сын был также Xg(a-). Описан фенотип Xg(a-) у дочерей, отцы которых были Xg(a+) (Sanger и соавт. [80], Tippett, Ellis [91]). В последнем варианте наследования нельзя исключить делецию локуса XG.


{kind=link}