Поиск по сайту
- Вы здесь:
-
Главная

-
Медицинские автомобили
-
book
-
Группы крови человека
- Система Kidd
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
Система Kidd
Антиген Jka, обнаружений в 1951 г. Allen и соавт. [3], назван по инициалам 6-го ребенка американской белой женщины, миссис Kidd, родившегося с проявлениями гемолитической болезни. Антитела анти-Лса реагировали с эритроцитами 77 % жителей г. Бостона (США).
Двумя годами позже (в Англии) Plaut и соавт. [78] нашли антитетичный антиген - Jkb.
Среди европеоидов 76,4 % содержали антиген Jka, 23,6 % - не содержали (Race, Sanger [82]). Эти данные позволили рассчитать частоту генов Jka и Jkb(0,5142 и 0,4858), а также частоту генотипов Jka/Jka, Jka/Jkbn Jkb/Jkb (0,2644; 0,4996 и 0,2360 соответственно). Указанные расчетные данные практически не отличались от фактических, полученных при обследовании 1051 канадской семьи сыворотками анти-Лса и анти-Лсь: ген Jka имел частоту 0,5162; Jkb -0,4838 (Chown и соавт. [7], Race и Sanger [82]).
По результатам ДНК-типирования 106 шведов, частота генов Jka и Jkbсоставила 0,53 и 0,47 соответственно (Irshaid и соавт. [35]).
Mourant и соавт. [62], Tills и соавт. [102] суммировали результаты популя-ционных исследований, выполненных различными авторами.
Ген Jka имеет частоту около 50 % среди жителей Европы, его частота выше (около 75 %) среди населения некоторых областей Африки и существенно ниже (20-30 %) в Азии (китайцы, японцы).
Гены Jka и Jkb передаются по наследству кодоминантно. При обследовании более чем 2000 семей европеоидов установлено, что ожидаемая и фактическая частота фенотипов совпадает (Race и Sanger [82]). Существование редкого молчащего аллеля Jk не оказывало влияния на результаты исследования.
Таблица 11.2
Распределение фенотипов по системе Kidd у представителей разных рас
|
Фенотип |
Частота фенотипа (%) среди |
||
|
европеоидов |
негроидов |
монголоидов |
|
|
Jk(a+b-) |
26,3 |
51,1 |
23,2 |
|
Jk(a-b+) |
23,4 |
8Д |
26,8 |
|
Jk(a+b+) |
50,3 |
40,8 |
49,1 |
|
Jk(a-b-) |
< 0,001 |
< 0,001 |
0,9 (жители Полинезии) |
Различие генов Jka и Jkb обусловлено перемещением G 838 А, ведущим к аминокислотной замене Asp 280 Asn на четвертой экстрацеллюлярной петле Jk-гликопротеина (Olives и соавт. [71], Reid, Lomas-Francis [83], Westhoff, Reid [107]).
Нуклеотидная замена G 838 А в аллеле Jka расположена перед сайтом рестрикции МпП., ее идентифицируют при генотипировании (Sidoux-Walter и соавт. [93]).
Для определения генотипа системы Kidd используют ПЦР с аллельспецифи-ческими праймерами (Irshaid и соавт. [37], Hessner и соавт. [30]). В одной из модификаций применена одноэтапная ПЦР с использованием прямого Jka- и обратного ^-специфических праймеров. Продукты ПЦР (Jka и Jkb) четко отличались друг от друга (Irshaid и соавт. [37]).
Обработка эритроцитов папаином, фицином, трипсином, химотрипсином и проназой усиливает реакции антител анти-1ка, анти-Лсь и анти-ЛсЗ. При феноти-пировании лиц по системе Kidd нередко прибегают к предварительной обработке эритроцитов бромелином (Issitt, Anstee [38]). Антигены Jka, Jkb и Jk3 не разрушаются сиалидазой и сульфгидрильными редуцентами (Daniels [10]).
Задолго до выделения гликопротеина Kidd, картирования и клонирования гена Ж было известно, что эритроциты, лишенные антигенов Kidd [Jk(a-b-)], не лизируются в 2М растворе мочевины, а эритроциты, содержащие антигены Kidd, лизируются указанным раствором. Уже тогда возникло предположение, что антигены Kidd участвуют в транспорте мочевины.
Используя нейлоновые мембраны с фиксированными к ним аффинно-очищенными IgG-антителами анти-Лса, анти-Лсь и анти-ЛсЗ, Sinor и соавт. [96] выделили из оболочки эритроцитов субстрат с мол. массой 45 кДа, несущий Jk-антигенную активность. Иммунопреципитат, выделенный с помощью антител анти-ЛсЗ из эритроцитов всех фенотипов Kidd, за исключением Jk(a-b-), представлял собой гликопротеин с мол. массой 46-60 кДа. После удаления N-гликана из гликопептида обработкой его N-гликаназой мол. масса уменьшилась до 36 кДа (Olives и соавт. [70]).
Электронная микроскопия эритроцитов, обработанных антителами IgG анти-Jkb, меченными ферритином, позволила установить, что эритроциты Jk(a-b+) несут около 14 тыс. антигенных участков Jkb на 1 клетку (Masouredis и соавт. [53]).
Maraiuzzu и соавт. [50] с помощью гидрофобных соединений ртути ингибировали транспорт мочевины и нашли, что количество участков, транспортирующих мочевину, т. е. участков, несущих антигены Jk, составляет около 32 тыс. на 1 эритроцит.
Olives и соавт. [72] исследовали кДНК эритробластов человека с помощью ПЦР с праймерами, сконструированными по аминокислотной последовательности кроличьего транспортера мочевины. Иммунопреципитацией антителами анти-Л<3 авторы выделили полипептид с мол. массой 36 кДа.
Методом иммуноблоттинга с кроличьими антителами был получен полипептид с мол. массой 46-60 кДа, который присутствовал в эритроцитах всех фенотипов Kidd, кроме Jk(a-b-) (Olives и соавт. [70]). Позднее было установлено, что ген HUT11 представляет собой аберрантный транскрипт или является артефактом клонирования (Sidoux- Walter и соавт. [93]). Другой ген - HUTU А, кодирующий ппотаминовую кислоту вместо лизина в положении 44 и 2 дипептида -валин и глицин - вместо 3 после позиции 227, производит гликопротеин Kidd и транспортер мочевины эритроцитов (Sidoux-Walter и соавт. [93], Irshaid и соавт. [37]). Продукт этого гена имеет мол. массу 43 кДа, содержит 389 аминокислот и на 63 % идентичен кроличьему транспортеру мочевины. Протеин содержит 10 трансмембранных доменов. Один из двух N-гликозилированных участков (Asn 211) расположен на третьем экстрацеллюлярном домене.
Ген Ж (SLC14A1) имеет величину 30 кб и состоит из 11 экзонов (Lucien и соавт. [48], Irshaid и соавт. [36]). Экзоны 1-3 и часть четвертого (табл. 11.1) представляют нетранслируемый У регион, экзоны 4-11 кодируют протеин.
Участок, инициирующий трансляцию, расположен на 335 пар выше стартового транслируемого кодона в экзоне 4. Область между нуклеотидами -837 и -336 содержит эритроидоспецифические участки транскрипции GATA-1 и SP1, а также ТАТА-бокс и инвертированный СААТ-бокс (Lucien и соавт. [48]).
Идентифицированы 2 эритроидных транскрипта величиной 4,4 и 2 кб. Последний (меньший из них) образуется за счет пропуска в считывании экзона 3 (Lucien и соавт [48]).Таблица 11.1
Организация гена JK
|
Экзон |
Количество пар оснований |
Позиция кодируемых аминокислот |
Размер интрона, кб |
|
|
* |
** |
|||
|
1 |
93 |
|
0,7 |
|
|
2 |
64 |
|
2,4 |
|
|
3 |
157 |
|
3,1 |
|
|
4 |
172 |
1-50 |
0,6 |
0,543 |
|
5 |
190 |
51-113 |
3,55 |
3,0 |
|
6 |
129 |
114-156 |
1,9 |
2,0 |
|
jjj 7 |
193 |
157-221 |
2,5 |
2,5 |
|
8 |
148 |
222-270 |
0,27 |
0,217 |
|
9 |
135 |
271-315 |
8,6 |
9,0 |
|
10 |
50 |
316-332 |
1,4 |
1,4 |
|
11 |
551 |
333-389 |
|
|
По Lucien и соавт. [48], **по Irshaid и соавт. [36].
Антигены системы Kidd (Кидд) - Jka и Jkb - являются продуктами аллель-ных генов. Антигенные различия Jka/Jkb обусловлены заменой Asp 280 Asn. Распределение их у представителей разных рас неодинаково. Антитела против антигенов Kidd представляют опасность в клиническом плане, поскольку вызывают замедленные посттрансфузионные реакции.
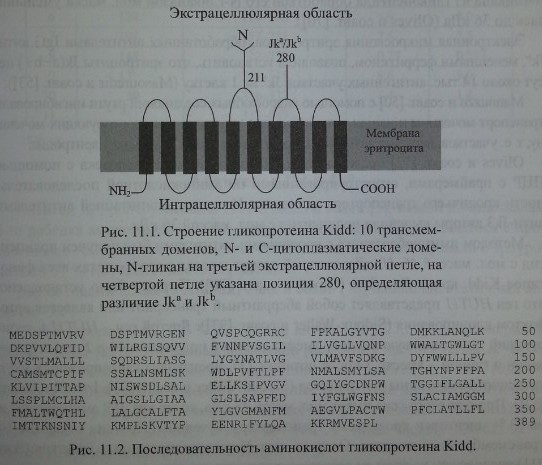
Антигены Kidd расположены на гликопротеине, имеющем 10 трансмембранных доменов (рис. 11.1). Определена его аминокислотная последовательность.
В системе Kidd известен нулевой фенотип Jk(a-b-), который чаще встречается у жителей Полинезии. На эритроцитах Jk(a-b-) отсутствуют антигены Jka и Jkb, а также антиген Jk3 (Jkab). Идентифицировано 5 мутаций, инактивирующих ген Ж.
Гликопротеин Kidd выполняет в клетке функцию транспортера мочевины. Генный локус Ж (SLC14A1) картирован на хромосоме 18 в позиции 18qll-ql2


{kind=link}