Поиск по сайту
- Вы здесь:
-
Главная

-
История
-
Великие лекарства
-
book
-
Группы крови человека
- Система Kidd
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
- Категория: Система Kidd
Антигены системы Kidd (Кидд) - Jka и Jkb - являются продуктами аллель-ных генов. Антигенные различия Jka/Jkb обусловлены заменой Asp 280 Asn. Распределение их у представителей разных рас неодинаково. Антитела против антигенов Kidd представляют опасность в клиническом плане, поскольку вызывают замедленные посттрансфузионные реакции.
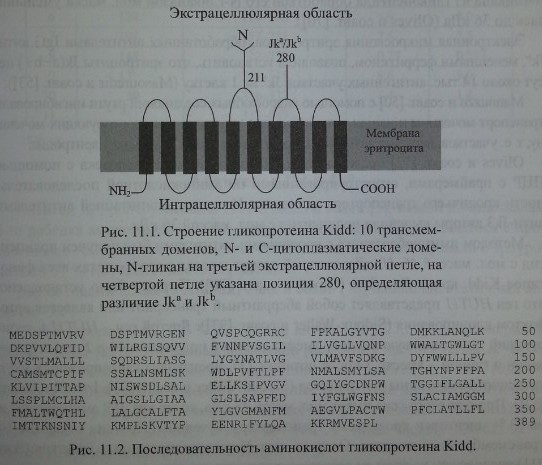
Антигены Kidd расположены на гликопротеине, имеющем 10 трансмембранных доменов (рис. 11.1). Определена его аминокислотная последовательность.
В системе Kidd известен нулевой фенотип Jk(a-b-), который чаще встречается у жителей Полинезии. На эритроцитах Jk(a-b-) отсутствуют антигены Jka и Jkb, а также антиген Jk3 (Jkab). Идентифицировано 5 мутаций, инактивирующих ген Ж.
Гликопротеин Kidd выполняет в клетке функцию транспортера мочевины. Генный локус Ж (SLC14A1) картирован на хромосоме 18 в позиции 18qll-ql2
- Категория: Система Kidd
Задолго до выделения гликопротеина Kidd, картирования и клонирования гена Ж было известно, что эритроциты, лишенные антигенов Kidd [Jk(a-b-)], не лизируются в 2М растворе мочевины, а эритроциты, содержащие антигены Kidd, лизируются указанным раствором. Уже тогда возникло предположение, что антигены Kidd участвуют в транспорте мочевины.
Используя нейлоновые мембраны с фиксированными к ним аффинно-очищенными IgG-антителами анти-Лса, анти-Лсь и анти-ЛсЗ, Sinor и соавт. [96] выделили из оболочки эритроцитов субстрат с мол. массой 45 кДа, несущий Jk-антигенную активность. Иммунопреципитат, выделенный с помощью антител анти-ЛсЗ из эритроцитов всех фенотипов Kidd, за исключением Jk(a-b-), представлял собой гликопротеин с мол. массой 46-60 кДа. После удаления N-гликана из гликопептида обработкой его N-гликаназой мол. масса уменьшилась до 36 кДа (Olives и соавт. [70]).
Электронная микроскопия эритроцитов, обработанных антителами IgG анти-Jkb, меченными ферритином, позволила установить, что эритроциты Jk(a-b+) несут около 14 тыс. антигенных участков Jkb на 1 клетку (Masouredis и соавт. [53]).
Maraiuzzu и соавт. [50] с помощью гидрофобных соединений ртути ингибировали транспорт мочевины и нашли, что количество участков, транспортирующих мочевину, т. е. участков, несущих антигены Jk, составляет около 32 тыс. на 1 эритроцит.
Olives и соавт. [72] исследовали кДНК эритробластов человека с помощью ПЦР с праймерами, сконструированными по аминокислотной последовательности кроличьего транспортера мочевины. Иммунопреципитацией антителами анти-Л<3 авторы выделили полипептид с мол. массой 36 кДа.
Методом иммуноблоттинга с кроличьими антителами был получен полипептид с мол. массой 46-60 кДа, который присутствовал в эритроцитах всех фенотипов Kidd, кроме Jk(a-b-) (Olives и соавт. [70]). Позднее было установлено, что ген HUT11 представляет собой аберрантный транскрипт или является артефактом клонирования (Sidoux- Walter и соавт. [93]). Другой ген - HUTU А, кодирующий ппотаминовую кислоту вместо лизина в положении 44 и 2 дипептида -валин и глицин - вместо 3 после позиции 227, производит гликопротеин Kidd и транспортер мочевины эритроцитов (Sidoux-Walter и соавт. [93], Irshaid и соавт. [37]). Продукт этого гена имеет мол. массу 43 кДа, содержит 389 аминокислот и на 63 % идентичен кроличьему транспортеру мочевины. Протеин содержит 10 трансмембранных доменов. Один из двух N-гликозилированных участков (Asn 211) расположен на третьем экстрацеллюлярном домене.
Ген Ж (SLC14A1) имеет величину 30 кб и состоит из 11 экзонов (Lucien и соавт. [48], Irshaid и соавт. [36]). Экзоны 1-3 и часть четвертого (табл. 11.1) представляют нетранслируемый У регион, экзоны 4-11 кодируют протеин.
Участок, инициирующий трансляцию, расположен на 335 пар выше стартового транслируемого кодона в экзоне 4. Область между нуклеотидами -837 и -336 содержит эритроидоспецифические участки транскрипции GATA-1 и SP1, а также ТАТА-бокс и инвертированный СААТ-бокс (Lucien и соавт. [48]).
Идентифицированы 2 эритроидных транскрипта величиной 4,4 и 2 кб. Последний (меньший из них) образуется за счет пропуска в считывании экзона 3 (Lucien и соавт [48]).Таблица 11.1
Организация гена JK
|
Экзон |
Количество пар оснований |
Позиция кодируемых аминокислот |
Размер интрона, кб |
|
|
* |
** |
|||
|
1 |
93 |
|
0,7 |
|
|
2 |
64 |
|
2,4 |
|
|
3 |
157 |
|
3,1 |
|
|
4 |
172 |
1-50 |
0,6 |
0,543 |
|
5 |
190 |
51-113 |
3,55 |
3,0 |
|
6 |
129 |
114-156 |
1,9 |
2,0 |
|
jjj 7 |
193 |
157-221 |
2,5 |
2,5 |
|
8 |
148 |
222-270 |
0,27 |
0,217 |
|
9 |
135 |
271-315 |
8,6 |
9,0 |
|
10 |
50 |
316-332 |
1,4 |
1,4 |
|
11 |
551 |
333-389 |
|
|
По Lucien и соавт. [48], **по Irshaid и соавт. [36].
- Категория: Система Kidd
Антитела анти-Jk* и анти-1кь
Вслед за первым описанием антител, найденных Allen и соавт. [3] в сыворотке крови мисс Kidd, появились другие сообщения о выявлении антител анти-Лса (М.А. Умнова и Т.М. Пискунова [1], Rosenfield и соавт. [85], Lindervall [46], Hunter и соавт. [34], Milne и соавт. [58], van der Hart, van Loghem [102], Greenwalt и соавт. [23]). Аналогично: после открытия Plaut и соавт. [78] антигена Jkb вскоре последовали другие публикации с описанием антител анти-1кь (Rosenfield и соавт. [84], Sanger и соавт. [90], van Loghem и соавт. [105], Geczy, Leslie [22]). Несмотря на то что в последующие десятилетия было найдено много других образцов сывороток с антителами системы Kidd, ни анти-1ка, ни тем более анти-Лсь не относят к широко распространенным антителам. Они часто сочетаются с антителами к другим групповым факторам эритроцитов, что свидетельствует о сравнительно невысоких иммуногенных свойствах антигенов системы Kidd. В литературе имеется единственное описание анти-Лса-антител естественного происхождения, обнаруженных у 9-месячных близнецов (Rumsey и соавт. [86]). В остальных случаях антитела системы Kidd находили у лиц, имевших переливания крови и беременности. В одном из наблюдений антитела анти-Лса появились во время первой беременности у женщины, не получавшей гемотрансфузии (Hunter и соавт. [34]). Описан случай образования анти-Лса после внутриматочной гемотрансфузии (Harrison, Popper [25]).
Антитела системы Kidd относятся к трудно выявляемым. Некоторые из них непосредственно агглютинируют нативные эритроциты, однако реакции слабо выражены (Allen и соавт. [3], Lindervall [46], Hunter и соавт. [34], Geczy, Leslie [22], Simmons и соавт. [95], Morgan и соавт. [60], Kronenberg и соавт. [42]). Для выявления антител системы Kidd наиболее подходящим является непрямой ан-тиглобулиновый метод Кумбса. Для обнаружения слабых антител эритроциты предварительно энзимируют.
Maynard и соавт. [55] описали антитела, выявляемые только в полибреновом тесте, которые вызвали гемолитическую посттрансфузионную реакцию.
Многие образцы анти-Лса-антител реагируют сильнее с эритроцитами Jk(a+b-), чем Jk(a+b+) (Race, Sanger [82], Crawford и соавт. [8]). При использовании эритроцитов Jk(a+b+), несущих только 1 дозу антигена Jka, слабые антитела указанной специфичности могут остаться не выявленными, что сопряжено с угрозой гемолитических посттрансфузионных реакций. В связи с этим панели для скрининга антиэритроцитарных антител комплектуют эритроцитами Jk(a+b-) и Лс(аЧ)+), но не Jk(a+b+), поскольку эффект дозы свойственен также и анти-Лсь-антителам.
Антитела системы Kidd представлены IgG или смесью IgG и IgM. Сыворотки, содержащие только IgM-антитела, встречаются редко (Polley и соавт. [80], Szymanski и соавт. [100]). В большинстве случаев антитела анти-Лса относятся к IgG3-cy6miaccy или являются смесью IgG3 и IgGl, иногда присутствуют только IgGl (Hardman, Beck [26]) или IgG2 (Szymanski и соавт. [100]). Из 6 образцов анти-Лсь-антител 4 относились к субклассу IgGl, 1 был представлен смесью IgGl и IgG3, 1 - IgGl и IgG4 (Hardman, Beck [26]). Примерно 40 % образцов антител Kidd обладает способностью связывать комплемент. Некоторые антитела могут быть выявлены лишь при условии, что антиглобулиновые реагенты содержат, помимо антиглобулиновых антител, преципитирующие антикомплементарные антитела (Mollison и соавт. [59], Yates и соавт. [ПО]). Способностью связывать комплемент обладали сыворотки, содержащие IgM компонент, IgG-антитела системы Kidd комплемент не связывают (Yates и соавт. [110]).
Некоторые антитела ингибировались ионами кальция (O'Brien и соавт. [65]).
Антитела анти-Jk3 вызывали тяжелые постгрансфузионные гемолитические реакции немедленного типа, в том числе с летальным исходом (Lindervall [46], Kronenberg и соавт. [42], Maynard и соавт. [55], Degnan, Rosenfield [12], Polesky, Bove [79]).
Описаны посттрансфузионные реакции замедленного типа. Они были тяжелыми, сопровождались олигурией, почечной недостаточностью со смертельным исходом (Mollison и соавт. [59], Pineda и соавт. [75], Ness и соавт. [64]).
Антитела анти-Лсь также вызывали тяжелые посттрансфузионные реакции (Morgan и соавт. [60], Holland, Wallerstein [31]).
Одной из особенностей посттрансфузионных реакций, вызванных Kidd-антителами, является быстрое снижение активности антител, в силу чего они могут быть не выявлены в последующих тестах на индивидуальную совместимость (Rosenfield и соавт. [85], Lindervall [46], Morgan и соавт. [60], Matson и соавт. [54]).
Pineda и соавт. [75, 76] подсчитали, что каждая третья замедленная пост-трансфузионная реакция обусловлена анти-Лса-антителами. По данным Чинь Суан Киэм и соавт. [2], проанализировавших в одном из вьетнамских госпиталей 63 случая гемолитических посттрансфузионных реакций замедленного типа, в 9 случаях выявлены антитела системы Kidd (5 анти-Jk3, 4 анти-Лсь).
Элиминация несовместимых эритроцитов в организме реципиента, содержащего антитела системы Kidd, происходит быстро (Maynard и соавт. [55], Cutbush, Mollison [9], Howard и соавт. [33]).
В реакции с монослоем моноцитов активность антител анти-Jk3 и анти-1кь усиливалась в присутствии комплемента (Nance и соавт. [63], Zupanska и соавт. [112]).
В 1 случае антитела анти-Jk3 вызвали гемолиз и снижение уровня гемоглобина у реципиента Jk(a+) после трансплантации костного мозга от сестры Jk(a-). В данном случае имела место реакция трансплантат против хозяина: антитела против эритроцитов реципиента вырабатывали пересаженные иммунокомпе-тентные клетки донора-сестры, которая, по-видимому, ранее была сенсибилизирована антигеном Jka. Лечение циклоспорином и метотрексатом устранило вын работку антител (Leo и соавт. [45]).
Антитела Kidd не вызывают клинически выраженной ГБН (Dorner и соавт. [13]). Описан всего один случай тяжелой ГБН, обусловленной антителами анти-Jk3 и сопровождающейся ядерной желтухой (Matson и соавт. [54]). Причины отсутствия ГБН, даже при высоких титрах антител Kidd, не выяснены.
Помимо аллоиммунных, найдены аутоиммунные антитела Kidd (van Loghem, van der Hart [106], Patten и соавт. [73], Strikas и соавт. [99], Sosler и соавт. [98], Sander и соавт. [87], Gandy и соавт. [21]), которые в ряде случаев вызывали аутоиммунную гемолитическую анемию. Один из больных, у которого, помимо аутоиммунной гемолитической анемии, развилась идиопатическая тром-боцитопеническая пурпура, был фенотипирован сначала как Jk(a-b+), однако позднее оказалось, что он относится к группе Jk(a+b+), что было подтверждено исследованием членов его семьи (Gandy и соавт. [21]).
Patten и соавт. [73] описали больного аутоиммунной гемолитической анемией, развившейся вследствие лечения альдометом. Больной имел фенотип Jk(a+), аутоантитела анти-Jk3 присутствовали как в сыворотке его крови, так и в элю-атах с эритроцитов. Прекращение приема лекарства привело к постепенному исчезновению аутоантител, клинические проявления гемолитической анемии также постепенно исчезали.
Аналогичный случай, сопровождавшийся острым гемолизом, описан у женщины Jk(a+b+), принимавшей хлорпропамид с целью лечения сахарного диабета (Sosler и соавт. [98]). В сыворотке ее крови имелись антитела анти-Jk3, которые реагировали с эритроцитами Jk(a+b+) в присутствии хлорпропамида или его аналогов. Гемолиз прекратился после отмены хлорпропамида. Не исключено, что субстанция Kidd, являясь транспортером азотистых шлаков, может иметь сродство к хлопропамиду, структура которого напоминает мочевину.
Аутоантитела анти-Jk3 находили у здоровых лиц (Holmes и соавт. [32], Issitt и соавт. [40]). Четыре образца таких антител реагировали с эритроцитами в присутствии парабенов (бутилового, этилового, метилового и пропилового эфиров /нгидроксилбензоата) и других ароматических соединений (Judd и соавт. [41], Halima и соавт.
Judd и соавт. [41] высказали предположение, что парабены способны вызвать обратимые структурные изменения антигена Jka, инициирующие образование аутоантител. Бензойные кольца парабенов взаимодействуют с аминокислотами гликопротеина Jk, при этом вовлеченными оказываются участки, формирующие эпитопы Jka.
Описан больной Jk(a-b+), перенесший нефрэктомию, с наличием хронической инфекции, вызванной протеем. У него имелся гемолиз, сыворотка содержала аутоантитела aHra-Jkb. Эритроциты Jk(b-), инкубированные с Proteus mirabilis, приобретали способность реагировать с антителами airra-Jkb (McGinnis и соавт. [57]).
В последние годы получены моноклональные антитела анти-Jk3 и анти-.Лсь класса IgM иммортализацией иммунных лимфоцитов с вирусом Эпштейна- Барр или гибридизацией их с клетками мышиной миеломы (Lecointre-Coatmelec и соавт. [44], Thompson и соавт. [101]). Полученные таким образом гетерогибридо-мы продуцировали агглютинирующие антитела анти-Jk3 и анти-Jk.
- Категория: Система Kidd
Антигены Jka и Jkb
Антиген Jka, обнаружений в 1951 г. Allen и соавт. [3], назван по инициалам 6-го ребенка американской белой женщины, миссис Kidd, родившегося с проявлениями гемолитической болезни. Антитела анти-Лса реагировали с эритроцитами 77 % жителей г. Бостона (США).
Двумя годами позже (в Англии) Plaut и соавт. [78] нашли антитетичный антиген - Jkb.
Среди европеоидов 76,4 % содержали антиген Jka, 23,6 % - не содержали (Race, Sanger [82]). Эти данные позволили рассчитать частоту генов Jka и Jkb(0,5142 и 0,4858), а также частоту генотипов Jka/Jka, Jka/Jkbn Jkb/Jkb (0,2644; 0,4996 и 0,2360 соответственно). Указанные расчетные данные практически не отличались от фактических, полученных при обследовании 1051 канадской семьи сыворотками анти-Лса и анти-Лсь: ген Jka имел частоту 0,5162; Jkb -0,4838 (Chown и соавт. [7], Race и Sanger [82]).
По результатам ДНК-типирования 106 шведов, частота генов Jka и Jkbсоставила 0,53 и 0,47 соответственно (Irshaid и соавт. [35]).
Mourant и соавт. [62], Tills и соавт. [102] суммировали результаты популя-ционных исследований, выполненных различными авторами.
Ген Jka имеет частоту около 50 % среди жителей Европы, его частота выше (около 75 %) среди населения некоторых областей Африки и существенно ниже (20-30 %) в Азии (китайцы, японцы).
Гены Jka и Jkb передаются по наследству кодоминантно. При обследовании более чем 2000 семей европеоидов установлено, что ожидаемая и фактическая частота фенотипов совпадает (Race и Sanger [82]). Существование редкого молчащего аллеля Jk не оказывало влияния на результаты исследования.
Таблица 11.2
Распределение фенотипов по системе Kidd у представителей разных рас
|
Фенотип |
Частота фенотипа (%) среди |
||
|
европеоидов |
негроидов |
монголоидов |
|
|
Jk(a+b-) |
26,3 |
51,1 |
23,2 |
|
Jk(a-b+) |
23,4 |
8Д |
26,8 |
|
Jk(a+b+) |
50,3 |
40,8 |
49,1 |
|
Jk(a-b-) |
< 0,001 |
< 0,001 |
0,9 (жители Полинезии) |
Различие генов Jka и Jkb обусловлено перемещением G 838 А, ведущим к аминокислотной замене Asp 280 Asn на четвертой экстрацеллюлярной петле Jk-гликопротеина (Olives и соавт. [71], Reid, Lomas-Francis [83], Westhoff, Reid [107]).
Нуклеотидная замена G 838 А в аллеле Jka расположена перед сайтом рестрикции МпП., ее идентифицируют при генотипировании (Sidoux-Walter и соавт. [93]).
Для определения генотипа системы Kidd используют ПЦР с аллельспецифи-ческими праймерами (Irshaid и соавт. [37], Hessner и соавт. [30]). В одной из модификаций применена одноэтапная ПЦР с использованием прямого Jka- и обратного ^-специфических праймеров. Продукты ПЦР (Jka и Jkb) четко отличались друг от друга (Irshaid и соавт. [37]).
Обработка эритроцитов папаином, фицином, трипсином, химотрипсином и проназой усиливает реакции антител анти-1ка, анти-Лсь и анти-ЛсЗ. При феноти-пировании лиц по системе Kidd нередко прибегают к предварительной обработке эритроцитов бромелином (Issitt, Anstee [38]). Антигены Jka, Jkb и Jk3 не разрушаются сиалидазой и сульфгидрильными редуцентами (Daniels [10]).
- Категория: Система Kidd
Нулевой фенотип Jk(a-b-), описанный в 1959 г. Pinkerton и соавт. [77], был обнаружен у жительницы Филиппин испано-китайского происхождения, имевшей 2 детей. После гемотрансфузии у нее развилась желтуха. Сыворотка ее крови реагировала со всеми образцами эритроцитов, кроме собственных Jk(a-b-). Адсорбция сыворотки эритроцитами Jk(a-b+) полностью устраняла активность антител, а адсорбция эритроцитами Jk(a+b-) - анти-1кь-активность не полностью. Элюаты реагировали с эритроцитами Jk(a+b-) и Jk(a-b+). Анализ результатов исследования позволил сделать вывод, что сыворотка женщины содержала антитела анти-1кь и ранее неизвестные несе-парируемые антитела анти-JkаЬ, или анти-1кЗ.
Фенотип Jk(a-b-) найден у жителей Полинезии (Irshaid и соавт. [36], Yokoyama и соавт. [Ill], Woodfield и соавт. [108], Kuczmarski и соавт. [43],
Heaton, McLoughlin [28], Henry, Woodfield [29], Sidoux-Walter и соавт. [92]),
Филиппин и Индонезии (Tills и соавт. [102], Marshall и соавт. [52], Pierse и соавт. [74]), китайцев (Lucien и соавт. [48], Day и соавт. [11]), японцев (Okubo и соавт. [67]), индусов (Tills и соавт. [102], Smart и соавт. [97]), бразильских индейцев (Tills и соавт. [102], Silber и соавт. [94]), американских негров (Oliver и соавт. [68]), жителей Туниса (Lucien и соавт. [47]), лиц белой расы, включая финнов (Irshaid и соавт. [36], Sidoux-Walter и соавт. [92]), Sareneva и соавт. [91]), французов (Habibi и соавт. [24]), швейцарцев и англичан (Irshaid и соавт. [35]).
Многие лица Jk(a-b-) были обнаружены в связи с наличием у них антител анти-ЛсЗ (Yokoyama и соавт. [Ill], Агсага и соавт. [4], Habibi и соавт. [24], Irshaid и соавт. [35], Kuczmarski и соавт. [43], Pierse и соавт. [73], Day и соавт. [11], Sareneva и соавт. [91]).
Чаще всего фенотип Jk(a-b-) выявляли у жителей Полинезии. Из 173 ООО полинезийских доноров 47 (0,27%) имели фенотип Jk:-1,-2,-3 (Henry, Woodfield [29]). Чаще всего указанный фенотип встречался среди племен Ниуэа и Тонга (1,4 и 1,2 % соответственно). Для поиска лиц Jk:-1,-2,-3 в других этнических группах использовали скрининговый тест на лизис эритроцитов в растворе мочевины и подтверждающее серологическое исследование. Среди 638 460 японских доноров г. Осака 14 имели фенотип Jk:-l,-2,-3, при этом 12 (0,002 %) относились к рецессивному типу (Okubo и соавт. [67]). Скрининговые исследования среди 13 817 жителей провинции Наталь в ЮАР выявили одного индивида Jk:-1,-2,-3 индийского происхождения (Smart и соавт. [97]). Среди 52 908 англичан и 120 тыс. жителей Новой Зеландии, преимущественно европейского происхождения, фенотип Jk:-1,-2,-3 не был найден (Heaton, McLoughlin [28], McDougall и соавт. [56]). Данная редкая группа была обнаружена у 24 (0,03 %) из 79 349 финнов (Sareneva и соавт. [91]).
Необычное наследование генов Jka и Jkb, свидетельствующее о существовании молчащего аллеля Jk, наблюдали в семье американцев, в которой был найден фенотип Lunull (Crawford и соавт. [8]), а также в афроамериканской семье (Behzad и соавт. [5]).
Установлено, что к возникновению фенотипа Jk:-1,-2,-3 может приводить гомозиготность по 5 различным мутациям (табл. 11.3):
- у жителей Полинезии идентифицирована мутация G —> А в инвариантном 3'-акцепторном участке сплайсинга в интроне 5 аллеля Jka. Последняя приводила к утрате экзона 6 в иРНК-транскриптах (Lucien и соавт. [48], Irshaid и соавт. [36]). Из 46 жителей Полинезии 8 были гетерозиготными по данной мутации. Аналогичная мутация выявлена также у китайцев, австралийцев, американцев и европейцев, имевших фенотип Jk(a-b-) (Sidoux-Walter и соавт. [92]);
- точковая мутация найдена у финнов: перемещение Т 871 С в аллеле Jkbприводило к аминокислотной замене Ser 291 Pro (Irshaid и соавт. [36], Sidoux-Walter и соавт. [92]);
- I у француза, имевшего фенотип Jk(a-b-), генная конверсия G > Т на участке сплайсинга в интроне 7 аллеля Jka способствовала исчезновению иРНК-транскрипта экзона 7 (Lucien и соавт. [48]);
- у 2 сестер англичанок и 1 тунисца, имевших фенотип Jk(a-b-), обнаружена делеция размером 1,6 кб в экзонах 4 и 5 аллеля ./fea(Irshaid и соавт, [35], Lucien и соавт. [47]);
- у 3 сестер из швейцарской семьи выявлена нонсенс-мутация в экзоне 7 аллеля Jka, трансформирующая кодон для тирозина в позиции 194 в стоп-кодон. Данная мутация приводила к синтезу укороченного полипептида, состоящего из 193 аминокислот (Irshaid и соавт. [35]).
Все перечисленные выше мутации выявлены с помощью ПЦР с соответствующими праймерами.
Ранее для выявления эритроцитов Jk(a-b-) использовали пробу Heaton и McLoughlin [28], предложенную ими в 1982 г. Метод основан на определении устойчивости эритроцитов к лизису в растворе мочевины.
Интересна история создания этого метода. У жителя Самоа, больного апла-стической анемией, автоматический счетчик клеток зарегистрировал повышенное содержание тромбоцитов. В счетчике с целью удаления эритроцитов, мешающих подсчету, использовали 2М раствор мочевины, в котором эритроциты обычно лизируются. Ошибочное завышение количества тромбоцитов произошло из-за высокой осмоустойчивости эритроцитов больного, оказавшегося Jk(a-b-), к мочевине. Не подвергшиеся лизису эритроциты и были приняты автоматическим счетчиком за тромбоциты. Эритроциты лиц, имеющих другие фенотипы Kidd, подвергаются лизису в 2М растворе мочевины в течение 1 мин, в то время как для лизиса эритроцитов Jk(a-b-) требуется не менее 30 мин. Проба на лизис с мочевиной оказалась весьма полезной для поиска эритроцитов редкого фенотипа Jk(a-b-) (Woodfield и соавт. [108], Henry, Woodfield [29], Okubo и соавт. [67], Smart и соавт. [97], Sareneva и соавт. [91], McDougall, McGregor [56]).
Эритроциты лиц, гетерозиготных по молчащему аллелю Jk, показали промежуточное время лизиса в растворе мочевины - между временем лизиса эритроцитов обычных фенотипов Kidd и временем лизиса эритроцитов Jk(a-b-) (Edwards-Moulds, Kasschau [16]).
Молекулярная основа нулевых фенотипов Kidd
|
Мутация |
Народы |
|
15 — 1 g > а; пропуск экзона 6 |
Французы |
|
17 + 1 g > t; пропуск экзона 6 |
Полинезийцы и китайцы |
|
Т 871 С в экзоне 9; Ser 293 Pro |
Финны |
|
С 582 G в экзоне 7; Туг 194 Stop, делеция экзонов 4 и 5 |
Швейцарцы |
|
Делеция экзонов 4 и 5 в геномной ДНК, экзонов 3-5 в кДНК |
Англичане |


{kind=link}