Поиск по сайту
- Вы здесь:
-
Главная

-
История
-
Великие лекарства
-
book
-
Группы крови человека
- Система Cartwright и система Xg
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
- Категория: Система Cartwright и система Xg
Система Cartwright (Картрайт) интересна тем, что носителями составляющих ее антигенов являются молекулы ацетилхолинэстеразы (АХЭ). Этот фермент участвует в передаче нервного импульса. Импульс передается на воспринимающие рецепторы очередного нейрона или мускульной клетки через синапс посредством образования ацетилхолина (АХ), который после проведения импульса разлагается АХЭ на холин и уксусную кислоту. Таким образом АХЭ выполняет роль биохимического реле, разделяющего нервные импульсы и одновременно контролирующего состояние системы связи. Острая токсичность фос-форорганических соединений является прямым следствием того, что они являются сильными ингибиторами АХЭ.
Ассоциационно-диссоциационная система АХ - АХЭ - АХ является основой иннервации всего организма, включая периферическую (проводниковую) и центральную нервную систему. Благодаря ей нервные импульсы поступают во все ткани организма.
В связи с этим можно высказать предположение о своеобразной иннервации циркулирующих клеток как в норме, так и в состоянии стресса. Стресс охватывает весь организм: и стационарные ткани, к которым подходят нервные окончания, и циркулирующие в кровотоке клетки, которых нервные окончания не достигают. Наличие АХЭ на эритроцитах, по-видимому, обеспечивает подведение к ним нервного импульса (доведение до эритроцитов стрессового сигнала). Организм как целостная система таким образом доводит сигнал стресса и мобилизационный импульс до всех тканей, стационарных и подвижных, обеспечивая ответную реакцию всего организма.
В литературе имеются указания на то, что ген ACHE (acetylcholinesterase) играет определенную роль в гемопоэзе. Аномалии хромосомы 7, где располагается этот ген, часто обнаруживают у пациентов с острым нелимфоцитарным лейкозом и миелодисплазией, и наиболее частые хромосомные нарушения при этих состояниях происходят в 7q22, участке гена ACHE (Кеге и соавт., 1989).
Lapidot-Lifson и соавт. (1989) связывали аномальный мегакариоцитопо-эз с нарушениями в локусе ACHE, вызванными химиотерапией, облучением или отравлением фосфорорганическими соединениями. Факт влияния АХЭ на мегакариоцитопоэз подтверждают результаты экспериментов на мышах (Patinkin и соавт., 1989).
- Категория: Система Cartwright и система Xg
Антигены Yta и Ytb
Система Cartwright представлена двумя антигенами: Yta и У1ь(табл. 13.1). Антитела анти-Yt8, открывающие часто встречающийся антиген Yta, обнаружены в 1956 г. Eaton и соавт. [12] при проведении пробы на индивидуальную совместимость. Через 8 лет Giles и Metaxas [16] описали антитетичный антиген Ytb, встречающийся относительно редко (с частотой Щ 8 %).
Таблица 13.1
Антигены системы Yt
|
Обозначение |
Характеристика |
||
|
традиционное |
ISBT |
Частота, % |
Молекулярная основа |
|
Yta [Yt(a+b-)] |
YT1 |
91,8 |
His353 |
|
Ytab [Yt(a+b+)] |
нет |
7,8 |
|
|
Ytb [Yt(a-b+)] |
YT2 |
0,3 |
Asn353 |
|
Yt(a-b-) |
нет |
|
|
Нулевой фенотип Yt(a-b-), как и молчащий ген, передаваемый по наследству, в системе Yt неизвестен.
Антигенные различия Yta/Ytb обусловлены заменой гастидина на аспарагин в позиции 353 (рис. 13.1). Локусы УТиАСНЕ (рис. 13.2) не зависят от генов других групп крови. Они расположены на хромосоме 7 в позиции 7q22.1 (Reid, Lomas-Francis [40]). В 1989-1991 гг. группа исследователей (Coghlan и соавт., Zelinski и соавт.) опубликовала данные о возможной сцепленности локусов YT и KEL. Последний, как вскоре выяснилось, так же как и УГ, расположен на хромосоме 7. Однако последующие исследования не подтвердили существование указанной сцепленности.
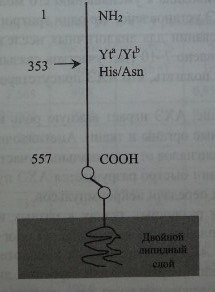
Строение гликопротеина, несущего антигены Yt.
- Категория: Система Cartwright и система Xg
Из 1568 доноров, обследованных Eaton и соавт. [12], Giles и соавт. [17] в Оксфорде, только 5 имели фенотип Yt(a-), остальные - Yt(a+). Таким образом, частота антигена Yta составила 99,8 %, гена Yta - 0,9559 (табл. 13.2). Установлено, что 8 % лиц белой расы имели фенотип Yt(b+). По результатам изучения частоты антигенов YtaH Ytb рассчитана частота генотипов: Yta/Yta(0,8966), Yta/Ytb (0,1006) и Ytb/Ytb (0,0028). Расчеты показали, что существование третьего аллеля в системе Cartwright вряд ли возможно
Распределение антигенов Yta и Ytbу различных народов
|
Популяция |
п |
Частота фенотипа, % |
Частота гена |
Источник |
||
|
Yt(a+) |
Yt(b+) |
Yta |
|
|||
|
Англичане |
2 568 |
99,8 |
— |
0,9559 |
0,0441 |
[12,17] |
|
Валлийцы (уэльсцы) |
29 802 |
99,9 |
— |
0,9761 |
0,0239 |
[15] |
|
Европейцы |
1399 |
— |
8,1 |
0,9587 |
0,0413 |
[17] |
|
Французы |
7 510 |
99,7 |
8,1 |
0,958 |
0,042 |
[42] |
|
Канадцы (белые) |
659 |
100,0 |
10,6 |
0,9469 |
0,0531 |
[28] |
|
Американские негры |
714 |
— |
8,4 |
0,9571 |
0,0429 |
[50] |
|
Евреи (израильские) |
2 549 |
98,6 |
21,3 |
0,8845 |
0,1154 |
[25] |
|
Арабы (израильские) |
85 |
97,6 |
23,5 |
0,8706 |
0,1294 |
[25] |
|
Друзы (израильские) |
77 |
97,4 |
26,0 |
0,8571 |
0,1429 |
[25] |
|
Японцы |
5 000 |
100,0 |
1,0000 |
0,0000 |
[34] |
|
Примечание: « - » - нет данных.
Обследование лиц различной национальности показало, что среди евреев, арабов и друзов Израиля антиген Ytb встречается относительно часто (табл. 13.2). В то же время этот фактор не был выявлен у японцев, жителей Таиланда, а также у инков и других коренных жителей Южной Америки (Nakajima и соавт. [34], Mourant и соавт. [33]).
Giles и соавт. [17] установили среди 1399 обследованных 113 (8,1 %) лиц Yt(b+). Salmon и соавт. [42] выявили 608 человек Yt(b+) среди 7510 французов (8,1 %). Lewis и соавт. [28] обследовали 659 канадцев, из них 589 были Yt(a+b~), 70 Yt(a+b+), не было ни одного Yt(a-b+). Wurzel и Haesler [50] протестировали эритроциты 714 американских негров и обнаружили, что 60 из них (8,4 %) были Yt(b+). Эти данные показывают, что частота Yt* и Ytb одинакова у негроидов и европеоидов.
Обращают на себя внимание две популяции, в которых частота антигенов Yta и Ytb отличается от таковой в других сравниваемых группах. Так, Nakajima и соавт. [34] протестировали 5 тыс. японских доноров, из них 70 были протестированы анти-У^-сывороткой. Все японцы были Yt(a+), ни одного индивида Yt(b+) не обнаружено. Возможно, для японцев характерен исключительно фенотип Yt(a+b-).
Отмечена более высокая, чем ожидалась, частота анти-У1:а-антител у израильтян. Когда появились тестовые анти-У1ь-реактивы, было установлено, что частота этого антигена у жителей Израиля существенно выше, чем в других географических зонах. Повышенная частота антигена Ytb в этом регионе привела к некоторому увеличению частоты фенотипа Yt(a-b+), что могло повлиять на частоту аллоиммунизации лиц Yt(a-b+). Из 1683 целенаправленно протестированных израильских евреев, арабов и друзов 26 имели фенотип Yt(a-b+). Иными словами, среди этих народов лиц Yt(a-b+) обнаруживают с частотой 1 на 65. Частота встречаемости этого фенотипа среди англичан, французов и негров составляет примерно 1 на 500, то есть почти в 10 раз меньше.
Результаты двух посемейных исследований свидетельствовали о кодоминантном наследовании генов Ytan Ytb в соответствии с законом Менделя. Данных о существовании молчащего аллеля не получено (Giles и соавт. [17], Lewis и соавт. [28]).
С антигенными различиями Yta/Ytb ассоциированы две нуклеотидные замены внутри гена ACHE. Одна из них, С 1057 А в экзоне 2, кодирует замену гисти-дина на аспарагин в положении 353. Другая мутация, С 1432 Т в экзоне 3, фе-нотипически не проявляется. Третья замена нуклеотидов, в экзоне 5, также не проявляла себя в зрелом протеине (Battels и соавт. [3]).
Исследование АХЭ электрического ската Torpedo californica, наиболее изученной формы этого фермента, показало, что лишь аминокислотная замена в позиции 353 приводит к формированию иммуногенного участка, способного стимулировать синтез специфических антител. Различия в антигенной структуре. АХЭ не влияют на ее активность (Masson и соавт. [30]).
Антиген Yta устойчив к действию трипсина, а-химотрипсин его разрушает. Обработка эритроцитов фицином и папаином дала разноречивые результаты, которые зависели от того, какими образцами анти-У^-антител проводили тестирование энзимированных эритроцитов (Eaton и соавт. [12], Vengelen-Tyler, Morel [48], Morton [32], Rouger и соавт. [41], Daniels [7]). В некоторых случаях обработка указанными протеолитическими ферментами усиливала реакцию. К действию сиалидазы антиген Yta не чувствителен (Rouger и соавт. [41], Daniels [7]).
Антигены Yta и Ytb разрушаются под воздействием дитиотрейтола, 2-ами-ноэтилизотиоурониумбромида (АЕТ), 2-меркаптоэтанола (Branch и соавт. [6], Levene, Harel [26], Shulman и соавт. [43]). Денатурация зависела от концентрации дитиотрейтола при обработке эритроцитов. Когда применяли 200 мМ препарата, как Yta, так и Ytb денатурировались. При использовании 50 и 100 мМ растворов дитиотрейтола активность антигена Yta снижалась пропорционально повышению концентрации раствора ДТТ, но не подавлялась полностью.
Shulman и соавт. [43] показали, что антиген Ytb денатурируется, когда эри-троцты обрабатывают 500 мМ раствором 2-меркаптоэтанола.
Уместно упомянуть, что применение АЕТ для создания искусственных эритроцитов Kellnull подверглось критике, поскольку этот препарат разрушает и другие, не относящиеся к системе Kell, антигены. Однако, когда Levene и Harel [26] обработали АЕТ эритроциты Yt(a+), денатурация антигена Yta не была такой полной, как денатурация антигенов системы Kell. Девять из 15 сывороток aHTH-Yta не реагировали с эритроцитами, обработанными АЕТ, другие 6 все же реагировали, хотя не так сильно, как с необработанными эритроцитами.
Антигены Yta и Ytb определяются на эритроцитах новорожденных: Ytb выражен слабо, в то время как экспрессия Yta не отличается от таковой у взрослых. Giles и соавт. [17] установили, что экспрессия антигена Ytb у новорожденных не изменена. Слабую экспрессию антигена Yta наблюдали у недоношенных (Eaton и соавт. [12], Ferguson и соавт. [14]); 8 из 10 образцов эритроцитов не реагировали с анти-У1ь-сывороткой, остальные 2 реагировали слабо (Gobel и соавт. [18]).
АХЭ присутствует в нервной и мышечной тканях и на эритроцитах (Taylor [46]). Данных о распределении этого фермента в других тканях мало. Антиген Yta не был выявлен на лимфоцитах, гранулоцитах и моноцитах при исследовании методом проточной цитофлюориметрии (Dunstan [11]).
- Категория: Система Cartwright и система Xg
Telen и соавт. [47] показали, что большинство образцов анти-У^-антител не реагирует с фракцией комплементчувствительных эритроцитов больных парок-сизмальной ночной гемоглобинурией (ПНГ), однако взаимодействует с фракцией эритроцитов, не чувствительных к комплементу. Установлено, что упомянутые комплементчувствительные эритроциты в значительной степени лишены глико-зилфосфатидилинозитола (ГФИ). У двух больных ПНГ комплементчувствительные эритроциты были Yt(b+). Эти результаты интерпретированы авторами как указание на то, что эпитопы YtaH Ytb несет ГФИ-ассоциированный гликопротеин.
В 1991 г. Spring и соавт. [45] показали, что антигены Cartwright на эритроцитах несет ГФИ-ассоциированный гликопротеин АХЭ (см. рис. 13.1). Путем иммунопреципитации антителами aHTH-Yta и анти-Yt15 выделен субстрат с мол. массой 72 кДа. По электрофоретической подвижности он был идентичен веществу, выделенному с помощью моноклональных антител к АХЭ эритроцитов. Субстрат обладал ацетилхолинэстеразной активностью, о чем свидетельствовали результаты экспериментов Petty [36] с иммобилизацией моно- и поликло-нальных антител анти-Yt3 и анти-Yt15.
АХЭ эритроцитов подвергается TV-гликозилированию. Как установили Spring и соавт. [45], обработка субстрата, выделенного посредством иммунопреципитации, N-гликаназой приводила к уменьшению его мол. массы с 72 до 63 кДа. С помощью МКА к АХЭ установлено, что один эритроцит несет 3-5 тыс. АХЭ-участков. При использовании для аналогичных исследований Fab-фрагментов указанных антител выявлено 7-10 тыс. участков связывания. Это дало основания Spring и соавт. [45] полагать, что АХЭ присутствует в мембране эритроцитов в форме димера.
Как указывалось выше, АХЭ играет важную роль в передаче нервных импульсов в исполнительные органы и ткани. Ацетилхолин обеспечивает передачу электроимпульсных сигналов от терминальных участков нервов в мышечные клетки. Затем ацетилхолин быстро разрушается АХЭ путем гидролиза для прекращения последующей передачи нейроимпульсов.
АХЭ присутствует в различных тканях в разных изоформах, обусловленных альтернативным сплайсингом гена ACHE (Taylor [46], Li и соавт. [29]). Фрагменты кДНК гена ACHE обнаружены в библиотеке генов из клеток фетальной мышечной ткани и нервной ткани взрослых лиц (Soreg и соавт. [44]). Полная кодирующая последовательность гена установлена Li и соавт. [29] при анализе космидной библиотеки генома человека.
Три экзона кодируют сигнальный пептид и N-терминальный участок, состоящий из 535 аминокислот (рис. 13.3). За счет альтернативного сплайсинга следующего экзона возникают структурные изменения в С-терминальном домене, позволяющие ГФИ присоединяться к эритроидным клеткам.
Аминокислотная последовательность протеина АХЭ
- Категория: Система Cartwright и система Xg
Антитела анти-Yt* и aHTH-Ytb
Несмотря на небольшое число лиц Yt(a-), способных к сенсибилизации антигеном Yta, антитела к этому антигену не являются редкостью и многократно описаны в литературе [1, 2,4, 5, 8-10,12,13,18,20,24,35]. Стандартизация 79 сывороток aHTH-Yta показала, что 57 из них были моноспецифическими, 22 содержали сопутствующие антитела другой специфичности (Eckrich и соавт. [13]).
Антиген Ytb в отличие от антигена Yta является более слабым иммуногеном. Антитела aHTH-Ytb встречаются редко и практически всегда обнаруживаются в сочетании с антителами другой специфичности (Giles и соавт. [16, 17], Ferguson и соавт. [14], Ikin и соавт. [21], Wurzel, Haesler [49], Levy и соавт. [27]). Так, согласно сводке Issitt и Anstee [22], в одной из сывороток aHTH-Ytb присутствовали анти-Fyb и анти-Вра (Bishop системы Diego), в другой - анти-С и анти-Hov (DI9 системы Diego), в третьей - анти-е, в четвертой - анти-К, в пятой -анти-Е и анти-Le3.
Следует отметить, что антитела к антигенам Yta и Ytb находили у беременных или реципиентов. Они ни разу не описаны как антитела естественного происхождения. Практически все образцы антител Yt были IgG с оптимумом реагирования при температуре 37 °С в непрямой антиглобулиновой пробе. Большинство образцов относилось к субклассу IgGl, нередко в сочетании с IgG4. Некоторые образцы были представлены только субклассом IgG4. Ни в одном из случаев антитела не относились к субклассу IgG3 (Vengelen-Tyler, Morel [48], Pierse и соавт. [37]). Некоторые образцы антител анти-Yt3 обладали способностью связывать комплемент (Bergvalds и соавт. [4]), в то время как у других такая способность отсутствовала (Gobel и соавт. [18], Ballas, Sherwood [2]).
Антитела системы Yt не описаны ни разу в качестве причины ГБН, хотя в ряде случаев были выявлены у женщин Yt(a-), родивших детей Yt(a+) (Wurzel, Haesler [50], Gobel и соавт. [18], Bergvalds и соавт. [4], Bettigole и соавт. [5], Lavallee и соавт. [24], Davey, Simkins [9]). У одной женщины Yt(b-), имевшей анти-У^-антитела, родился ребенок Yt(b+) без признаков ГБН (Ferguson и соавт. [14]). Антитела системы Yt считаются клинически значимыми в практике трансфузиологии. Антитела анти-Yt3 вызвали гемолитическую посттрансфузи-онную реакцию с летальным исходом у больного серповидно-клеточной анемией (Reed и соавт. [39]). Описаны посттрансфузионные реакции немедленного типа, обусловленные антителами анти-Yt3 (Hadley и соавт. [19]). Вместе с тем у многих реципиентов, имевших анти-У^-антитела и получавших трансфузии эритроцитов Yt(a+), признаков несовместимости не наблюдалось (Dobbs и соавт. [10], Eckrich и соавт. [13]). В одном случае антитела анти-Yt3 не вызвали посттрансфузионной реакции, несмотря на то, что в тестах in vitro проявляли гемолитические свойства, свидетельствовавшие об их трансфузионной опасности (AuBuchon и соавт. [1]). Из 18 реципиентов, имевших анти-У1а-антитела, только у 3 наблюдалось уменьшение продолжительности жизни перелитых эритроцитов (Eckrich и соавт. [13]). По данным других исследователей, длительность персистенции несовместимых эритроцитов в кровотоке реципиентов варьировала в широких пределах (Gobel и соавт. [18], Bettigole и соавт. [5], Dobbs и соавт. [10], Ballas, Sherwood [2], Davey, Simkins [9], Nance и соавт. [35], Kakaiya и соавт. [23]). Лишь в отдельных случаях имелись основания полагать, что перелитые эритроциты будут быстро элиминированы антителами из кровотока (Gobel и соавт. [18], Bettigole и соавт. [5], Ballas, Sherwood [2]). В исследованиях с использованием клеток, фагоцитирующих эритроциты, сенсибилизированные антителами, также получены противоречивые данные (Gobel и соавт. [18], AuBuchon и соавт. [1], Eckrich и соавт. [13], Levy и соавт. [27], Pierse и соавт. [37], Hadley и соавт. [19], Kakaiya и соавт. [23]). В связи с этим представляется очевидным, что каждый образец антител требует индивидуального исследования, по результатам которого можно прогнозировать степень несовместимости перелитых эритроцитов и, соответственно, безопасность и лечебную эффективность планируемой трансфузии.
При отсутствии совместимых эритроцитов Yt(a-) реципиенту, имеющему анти-У1а-антитела, переливают эритроциты Yt(a+). При этом целесообразно отобрать эритроциты, дающие менее сильную реакцию in vitro с сывороткой реципиента.
Описаны анти-У^-антитела у реципиента Yt(a+). Эти антитела реагировали с эритроцитами Yt(a+) неродственных лиц и матери реципиента, но не реагировали с эритроцитами самого реципиента и его Yt(a+) отца (Mazzi и соавт. [31]). Указанное наблюдение нельзя рассматривать как доказательство существования парциальных вариантов антигена Yta, поскольку анти-У1;а-антитела, появившиеся у реципиента на 9-й день после переливания 5 доз эритроцитов Yt(a+), через несколько месяцев исчезли.


{kind=link}