Вещества, вызывающие агглютинацию А-, В- или Н-положительных эритроцитов человека, были найдены во многих растениях, водорослях, грибах, а также в тканях некоторых червей, улиток, икре лососевых рыб, сыворотке крови угрей. Они были названы лектинами (табл. 3.6). Установлено, что они взаимодействуют с терминальными моносахаридными группировками. Лектины из Dolichos biflorus и Ulex europaeus используют для дифференцировки подгрупп А в исследовательских целях (П.Н. Косяков [35], Прокоп, Геллер [56], Daniels [87], Issitt, Anstee [122], Sringarm и соавт. [201]), в рутинной практике определения групп крови АВО их не применяют.
Таблица 3.6
Лектины с анти-А-, анти-В-, анти-AB- и анти-Н-активностью
|
Источник получения |
Специфичность |
|
|
Dolichos biflorus |
Семена |
Анти-А |
|
Phaseolus limensis |
To же |
Анти-А |
|
Phaseolus lunatus |
II |
То же |
|
Helix pomatia |
Печень |
II |
|
Helix hortensis |
To же |
II |
|
Fomes fometarius |
Мицелий |
АнпьВ |
|
Ptilota plumosa |
Органеллы |
То же |
|
Salmo salar |
Икра |
)1 |
|
Sophora japonica |
Семена |
Анти-АВ |
|
Phlomis Jructosa |
То же |
То же |
|
Bandeiraea simplicifolia |
II |
II |
|
Ulex europaeus |
II |
Анти-Н |
|
Lotus tetragonolous |
11 |
Анти-Н (HI) |
|
Anguilla anguilla |
Анти-Н (HI) |
|
|
Cystisus sessifolius |
Семена |
Анти-Н |
|
Laburnum alpinum |
То же |
То же |
Антиген С
Существование антигена С в системе АВО можно считать доказанным. Он содержится в эритроцитах А(П), В(Ш) и AB(IV). В эритроцитах 0(1) антиген С отсутствует.
Основанием для такого утверждения служат эксперименты Wiener и соавт. [213, 224, 228-230], показавшие, что сыворотки Оар(1), помимо а- и Р-агтлютининов, реагирующих с эритроцитами А и В, содержат еще одну разновидность естественных групповых антител - С-агтлютинины (по Винеру анти-С- антитела), которые реагируют с эритроцитами А и В перекрестно
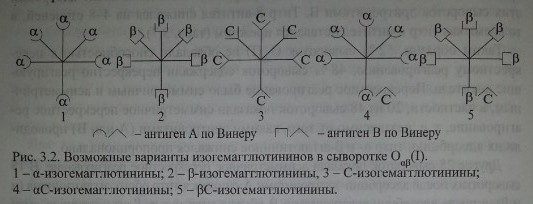
На наш взгляд, перекрестное реагирование обусловлено не ар-агглютининами, как полагала Dodd и соавт., а антителами аС, PC, что в большей мере согласуется с концепцией Винера, а также полученными нами данными. На рис. 3.2 представлена предполагаемая модель этих антител.
Представленная структурная модель изогемагглютининов позволяет объяснить характер перекрестного реагирования сывороток Оар(1). Сыворотки, не вызывающие перекрестных реакций, содержат только а- и (3-антитела (рис. 3.3, п. 1). Сыворотки, вызывающие симметричные перекрестные реакции, содержат, помимо а- и р-антител, антитела аС и PC или анти-С-антитела (рис. 3.3, п. 4). Сыворотки, вызывающие асимметричные перекрестные реакции, содержат а, р и одну из разновидностей антител - аС или PC (рис. 3.3, п. 2, 3).
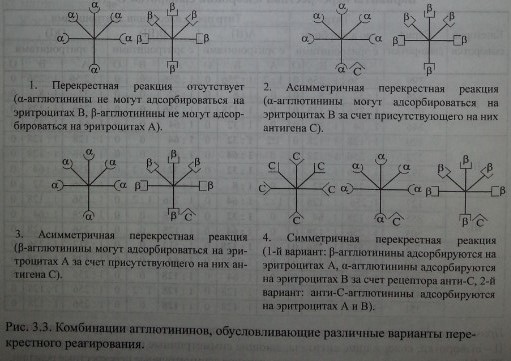
Комбинации агглютининов, обусловливающие различные варианты перекрестного реагирования.
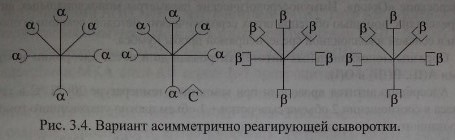
Вариант асимметрично реагирующей сыворотки.
Согласно схеме сыворотка содержит 50 % а-агглютининов, 75 % - р. Валентность анти-С реагирует как а и р. При адсорбции такой сыворотки эритроцитами А а-агглютинины будут удалены полностью. Иными словами, из сыворотки будет удалено 50 % антител. При адсорбции сыворотки эритроцитами В будет удалено 75 % антител (50 % Р+25 % а) за счет рецептора анти-С. В случае, если рецептор анти-С связан не с а-, а с р-агглютининами, асимметричное реагирование проявят эритроциты А, унося на своей поверхности 25 % Р-агглютининов.
Как далее нами показано, перекрестно реагирующие антитела так же, как иммунные антитела АВО и Rh, относятся к классу IgG, устойчивы к унитиолу, лучше реагируют с отмытыми, чем неотмытыми эритроцитами, в отличие от иммунных (термо- сгабильных) антител являются термолабильными (утрачивают активность после прогревания сыворотки при 70 °С в течение 10 мин). В противоположность классическим иммунным антителам IgG, которые адсорбируются на эритроцитах, но не вызывают их агглютинации, перекрестно реагирующие антитела непосредственно агглютинируют эритроциты в солевой среде. В низкой концентрации они утрачивают способность агглютинировать эритроциты, однако сенсибилизируют их подобно неполным антителам и хорошо выявляются в непрямой антиглобулиновой пробе Кумбса.
Сыворотки лиц 0^(1), очевидно, содержат несколько типов групповых антител: изогемагглютинины аир IgM, аир IgG, аС и рС IgG* анти-С IgG, иммунные аир IgM, иммунные а и Р IgG, что закономерно обусловливает перекрестные реакции с различными групповыми антигенами.
Важным аргументом, подтверждающим концепцию Винера о существовании антигена С, явились эксперименты по получению специфических антисьгворотк. В частности, при иммунизации мышей эритроцитами А(П) и В(Ш) нами [21] были получены моноклональные антитела со специфичностью анти-АВ (МКА-АВ). Материал и методы. Трехмесячным самкам мышей BALB/c вводили по 0,1 мл отмытых 0,9% NaCl эритроцитов А(П) или В(Ш) внутрибрюшинно дважды с интервалом в 7 дней. Третью инъекцию производили внутривенно за 5 дней до слияния спле- ноцитов с клетками миеломы NS-1. Слияние осуществляли по стандартной методике с использованием 45% полиэтиленгликоля-1500 «ЬоЬа» и 5% диметилсульфоксида.
Эффективность гибридизации составляла 10-5. Клонировали гибридные клетки в среде HAT без фидера. Антителопродуцирующие клоны отбирали с помощью реакции аг
в 96-луночных планшетах с отмытыми эритроцитами 0(1), А(П) и В(Щ), Класс моноклональных антител устанавливали по флюоресценции сенсибилизи-IgG, нейтрализовались группоспецифическими субстанциями, существенно отличает их от классических иммунных антител и позволяет выделить их в особую группу - иммунные IgG-антитела термолабильные легко нейтрализуемые. К этой же группе могут быть отнесены перекрестно реагирующие антитела лиц 0(1), имеющие идентичную иммуносерологическую характеристику - IgG термолабильные. Их происхождение (иммунное или естественное) остается неясным. Не исключено, что они так же, как МКА анти-АВ, являются иммунными.
Обработка эритроцитов В(Ш) протеолитическими ферментами, специфически разрушающими антиген В, лишала эритроциты групповых свойств. После такой обработки эритроциты В(Ш) в отличие от обработанных глутаровым альдегидом приобретали свойства эритроцитов 0(1) группы и не агглютинировались сыворотками анти-В. Одновременно исчезала их способность взаимодействовать с перекрестно реагирующими антителами МКА анти-АВ, а также перекрестно реагирующими антителами лиц 0(1).
По характеру реагирования полученные МКА анти-АВ вряд ли можно отнести к анти-А и анти-В. По-видимому, это одно, маскирующееся под анти-А и анти-В антитело, реагирующее с общей для эритроцитов А(П) и В(Ш) антигенной детерминантой. По своим свойствам эти антитела близки к перекрестно реагирующим антителам сывороток лиц 0(1), описанным выше. Сам факт получения антител анти-АВ в ответ на иммунизацию эритроцитами как А, так и В служит веским аргументом в пользу того, что оба антигена наряду с существенными серологическими различиями имеют идентичный эпитоп С, в равной мере являющийся сильным иммуногеном.
Таким образом, имеются все основания полагать, что групповая система АВО представлена не двумя агглютиногенами - А, В и двумя агглютининами - а, Р, а тремя агглютиногенами - А, В, С и тремя агглютининами - а, р и С.
Антиген С выявляют с помощью перекрестно реагирующих сывороток лиц Оар(1) [184, 229]. Именно фракция перекрестно реагирующих агглютининов представляет собой антитела анти-С. Последние можно удалить из сыворотки указанных лиц адсорбцией как А, так и В эритроцитами. При рутинном исследовании антиген С маскируется присутствием антигена А и/или В. Антитела анти-С в обычных методах исследования проявляют себя подобно несепари- руемой комбинации а- и Р-агглютининов. У лиц, имеющих группу крови А и В, агглютинины анти-С отсутствуют.
Не исключено, что антитела анти-С могут встречаться в чистом виде. Однако отличить их от анти-А и анти-В обычными методами не представляется возможным. У плодов 0(1) от матерей 0(1) находят антитела со свойствами анти-С, у плодов А(Н) и В(Ш) от матерей А(Н) и В(Ш) таких антител не находят.
Owen (цит. по Wiener [229]) получил чистую фракцию анти-С-агглютининов адсорбцией сывороток 0^(1) эритроцитами опоссума и кролика, которые, содержат парциальный антиген В и, по-видимому, не содержат антигена С. После такой обработки оставшиеся в сыворотке антитела анти-С реагировали с эритроцитами А и В, и их можно было удалить эритроцитами А или В в ад- <юрбционной пробе. Однако для окончательного доказательства того, что С-, о- и Р-агглютинины являются самостоятельными независимыми друг от друга антителами, не достает эксперимента, в котором антигены А и В были бы разрушены ферментативно (не реагировали бы с сыворотками анти-А и анти-В), но при этом сохранили бы антиген С (продолжали бы агглютинироваться анти-С- сывороткой или элюатом аР). Только в том случае, если эритроциты не агглютинируются а и р, но продолжают агглютинироваться анти-С-антителами, можно не только с достаточной степенью уверенности констатировать существование антигена С, но и с помощью такого реактива изучить закономерности наследования этого антигена, возможные его разновидности и комбинации. В наших экспериментах ферментативное разрушение В-антигена приводило одновременно к утрате их способности реагировать с МКА анти-АВ. Иными слова ми разрушение В-антигена влекло за собой разрушение антигена С. Однако, не смотря на отрицательный результат этого эксперимента, полученные нами данные в целом со всей очевидностью свидетельствуют о наличии третьего антигена в системе АВО, существование которого блестяще предсказал Wiener. глютинации

