К13-эффект
К13-эффект правильнее называть эффектом отсутствия гена К13, поскольку гаплотип К13-, а не гаплотип К13, содержит ген, подавляющий экспрессию других антигенов системы Kell и napa-Kell (см. К13).
Marsh и соавт. [262], впервые обнаружившие человека с фенотипом К13-, вначале полагали, что низкая экспрессия антигенов k, Kpb, Jsb, Ku и К12 на эритроцитах этого человека связана с его гетерозиготностью по гену К°. Эритроциты пробанда сильно реагировали с сывороткой анти-Кх, что свидетельствовало о наличии у этого человека гаплотипа К°. Генотип пробанда соответствовал формуле К13-/К13-К. Однако, как показали Kaita и соавт. [220], гетерозиготность по гену К° не могла быть причиной депрессии других Kell-антигенов, поскольку люди, являющиеся генетически K°/kKpbJsb, нормально экспрессируют антигены k, Крь и Jsb. Таким образом, Marsh и соавт. [262] оставалось сделать единственно правильный вывод относительно ослабления антигенов Kell у лиц К13-, а именно: низкая экспрессия Kell у лиц К13- обусловлена антитетичным партнером К13, который, будучи в гомозиготной форме и в позиции цис к другим генам KEL, кодирует более слабую, чем в норме, экспрессию антигенов Kell.
У людей К13- слабо выражены антигены К18 (Barrasso [98]), К19 и TOU (Sabo и соавт. [337]).
Вполне вероятно, что случаи ослабления Kell-антигенов, наблюдаемые многими авторами, связаны с гетерозиготностью по К13-, однако это пока невозможно доказать, поскольку отсутствуют сыворотки против указанного гипотетического аллеля К13.
Можно также предположить, что К13-эффект никак не связан с антигеном К13, а является проявлением пока неизвестного генетического процесса, приводящего к уменьшению синтеза антигенов Kell, в том числе антигена К13, который в этом случае не производится вовсе и его экспрессия соответствует 0.
Ожидаемые, но не встречающиеся гаплотипы KEL
Как известно, групповые антигены крови наследуются кодоминантно и независимо друг от друга. Однако существует много исключений, которые ставят под сомнение непреложность этого правила и еще раз подчеркивают огромное разнообразие не только форм групповых антигенов крови, но и способов их наследственной передачи. Приведем некоторые примеры:
- частичное доминирование (эпистазия) D над С в позиции цис. В серологических реакциях это проявляется следующим образом: эритроциты лиц CDe/cde (С и D цис) медленнее агглютинируются сывороткой анти-С и агглютинация выражена слабее, чем при тестировании эритроцитов лиц Cde/cDe (CnD транс);
- ослабление выраженности антигена D (Du) под действием гена С, расположенного по отношению к гену D в позиции транс. Этот феномен проявляется в том, что эритроциты лиц CDe/cde реагируют с сывороткой анти-D значительно сильнее, чем эритроциты лиц Cde/cDe;
- Кра-эффект - подавление синтеза антигенов Kell геном Кра; -К13-эффект - подавление синтеза антигенов Kell неизвестным геном, присутствующим в гаплотипе К13-.
Следует упомянуть еще об одном феномене - неравновесном сцеплении, которое проявляется в том, что антигены двух групповых систем предпочтительно наследуются вместе. Например, у людей, имеющих группу крови М, антиген S встречается в 2 раза чаще, чем у людей, имеющих группу крови N. Среди лиц М+ 72 % являются S+, среди N+ - 31 % S+. Считается, что ген S близко расположен к гену М, поэтому чаще наследуются вместе с ним, чем с геном N. Неравновесное сцепление генов Ми S по сравнению cNnSпослужило основанием полагать, что антигены MN и Ss представляют собой тесно связанную антигенную систему MNS.
В наследственной передаче антигенов Kell также проявляются свои особенности. Помимо Кра- и К13-эффекта существует еще одна необычная генетическая закономерность, а именно отсутствие Kell-гаплотипов, которые по логике вещей должны присутствовать. Речь идет о гаплотипах, кодирующих два редких антигена Kell и более, например: КраК, KJsa, KpaJsa, ККр° и др. Такие гаплотипы до сих пор не обнаружены.
Фенотипы, несущие два редких антигена, встречаются, например:
K+k+Kp(a+b+), K+k+Js(a+b+), K+k+Ul(a+)Kp(a-b+). Однако во всех случаях семейных исследований [154, 249, 251, 252, 288, 357, 402] установлено, что лица с фенотипом K+k+Kp(a+b+) генетически являются KKpb/kKpa, лица с фенотипом K+k+Js(a+b+) имеют генотип KJsb/kJsa, а лица с фенотипом K+k+Ul(a+)Kp(a-b+)- генотип KKpb/kUla.
Возникает вопрос: почему гены К и Кра, К и Jsa9 К и UIa, а также гены других редких Kell-антигенов не наследуются по два и более в одном гаплотипе?
До настоящего времени стройной генетической концепции, объясняющей это явление, не предложено. Некоторые авторы разделяют точку зрения Chown о том, что kKpbJsb представляет собой эволюционно более ранний гаплотип, который в результате единичных мутаций трансформировался в гаплотипы KKpbJsb, kKpaJsb, kKpcJsb и kKpbJsa. Далее логика нарушается, поскольку остальные 5 редких генов: Ula, Wka, К23, К24 и К25 - не имеют аллелей. Согласно предположению Chown, гаплотипы ККра, KJsa и KpaJsa могли сформироваться вследствие двойных мутаций, вероятность которых ничтожно мала в связи с редкостью генов К, Кра, Крс, Jsanjsp. Однако, как полагают Issitt и Anstee [204], с момента открытия редких антигенов Kell выполнено огромное количество исследований, и если бы такие двойные мутации (мутации предыдущих мутаций) существовали, они были бы обнаружены.
Не лишена оснований и другая точка зрения, что антигены системы Kell кодируются 10 гаплотипами (рис. 5.4). Один из них - общий (частый), 9 других -редкие. Первый гаплотип (общий) состоит из гена R- (R-минус), кодирующего все антигены Kell, за исключением редких. Другие 9 гаплотипов представлены одним из редких генов, которые кодируют 1 редкий и все остальные часто встречающиеся антигены.
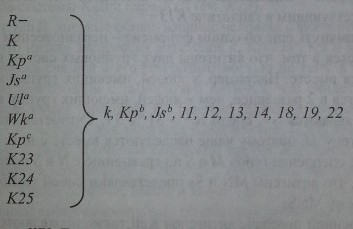
Гаплотипы KEL. Гаплотип 7?- производит общие Kell-антигены: k, Kpb, Jsb, 11, 12, 13,14,18,19 и 22; гаплотип К- антиген К и общие Kell-антигены; гаплотип Кра - антиген Краи общие Kell-антигены и т. д.
В соответствии с этой схемой гаплотипы, содержащие 2 редких антигена и более, исключены. Остается непонятным, почему в локусе KEL отсутствуют генные конверсии и другие формы обмена генетическим материалом между гомологичными хромосомами, как это имеет место в системе резус. По-видимому, в этом проявляется своеобразие рассматриваемого генного локуса.
Итак, подавляющее большинство людей гомозиготны по гаплотипу ШВ и соответственно содержат весь набор общих Kell-антигенов: k Kpb Jsb 11 12 13 14