Серологически выявляемая М-антигенная активность обусловлена присутствием серина в позиции 1 и глицина в позиции 5 на экстрацеллюлярном домене гликофорина A (Dahr и соавт. [55]). На N-положительных клетках в этих позициях находятся лейцин и глютаминовая кислота соответственно (рис. 6.1). В экспрессии антигенов важную роль играют О-гликаны и сиаловые кислоты. Позиции 2-4 пептидной цепи гликофорина А гликозилированы (Dahr и соавт. [55], Issitt, Anstee [113], Schenkel-Brunner [223]).
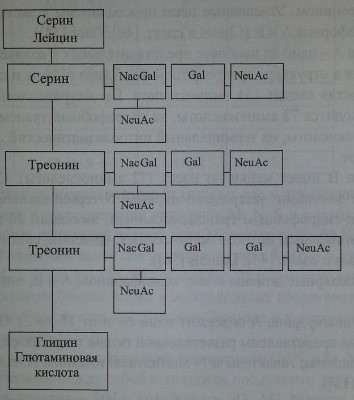
Строение терминальных участков гликофорина А.
Нередко антитела анти-М и анти-N обнаруживают у лиц, не имевших беременностей и гемотрансфузии (Garratty [78], Као и соавт. [128]). Такие антитела естественные, часто представляют собой IgM и реагируют при комнатной температуре как агглютинины. Описаны IgG-антитела той же специфичности (Garratty [78], Issitt, Anstee [113]). Многим антителам, особенно анти-М, свойствен эффект дозы. Эритроциты гомозигот М/М и N/N реагируют с антителами анти-М и анти-N сильнее, чем эритроциты гетерозигот. Большинство образцов указанных антител не обладает способностью активировать комплемент. Считается, что антитела анти-М и анти-N, относящиеся к Холодовым, не имеют клинического значения, поскольку не способны вызвать гемолиз эритроцитов in vivo (Garratty [78], Issitt, Anstee [113]). Дщучаи ||БН (Duguid и соавт. [65], Stone и соавт. [234], Teleschi и соавт 46]) й трансфузионные реакции (Alperin и соавт. [13], Ballas и соавт. 20], Furlong и соавт. [73], Sancho и соавт. [217]), обусловленные указанными антителами, редки.
В отечественной литературе имеется 2 сообщения об аллоиммунизации антигеном М в результате беременности. Выраженных клинических проявлений ГБН при этом не наблюдали (А.А. Михайлова, ТА. Ичаловская [6], А.Е. Скудицкий [8]).
Антитела анти-М обнаруживали у детей и взрослых, перенесших бактериальные инфекции (Као и соавт. [128]). Антитела выявляли у беременных, дети которых, как позже выяснилось, не имели антигена М. Анти-М-антитела иногда сопутствовали другим антителам (Rh, Kell, Kidd, Duffy) у гематологических больных, получивших многочисленные переливания компонентов крови (А. Майский, Л. Кучера [5], Gales и соавт. [77]).
Помимо естественных и аллоиммунных антител описаны аутоантитела анти-М и анти-N (Chapman и соавт. [39], Garratty и соавт. [79], Immel и соавт. [ПО], Sacher и соавт. [215]). В некоторых случаях течение аутоиммунной гемолитической анемии, обусловленной этими антителами, было тяжелым (Garratty и соавт. [79], Immel и соавт. [ПО]).
Различают рН-зависимые и глюкозозависимые антитела анти-М и анти-N (Beattie и соавт. [22], Reid и соавт. [200]). В отличие от обычных антител, для которых кислотность среды не столь существенна, рН-зависимые анти-М и анти-К-антитела наиболее активно реагируют с эритроцитами, несущими соответствующий антиген, при значении рН 6,5. Глюкозозависимые антитела проявляют свое действие лишь с эритроцитами, инкубированными в растворе глюкозы. Установлено, что активность некоторых образцов антител анти-М и анти-N ингибируется растворами мальтозы и маннита, но остается неизмененной в присутствии фруктозы, рибозы, галактозы, лактозы и сахарозы. Такие антитела выявляли главным образом у больных сахарным диабетом.
У больных почечной недостаточностью, находившихся на гемодиализе, обнаруживали особые антитела анти-N, реагирующие с эритроцитами, обработанными формальдегидом (Dzik и соавт. [66], Howell и соавт. [96], McLeish и соавт. [157], Sandler и соавт. [218], White и соавт. [259]). Такие антитела были обозначены как aHra-Nform. Дальнейшие исследования показали, что anra-Nf -антитела образовались в результате модификации мембраны эритроцитов больных следами формальдегида в гемодиализной системе (стерилизацию систем для гемодиализа проводили с использованием формальдегида).
AHTH-N-подобную специфичность имеют лектины растительного происхождения - Vicia graminea (А.К. Туманов, В.В. Томилин [9], Prigent [190]).
К настоящему времени получено большое количество образцов моноклональных антител анти-М и анти-N (Lisowska [149], Reid и соавт. [201]). Последние с успехом используют для фенотипирования эритроцитов, однако некоторые из них, в частности анти-М, не строго специфичны и в высоких концентрациях реагируют с эритроцитами M-N+, т. е. диспецифичны. Для придания им анти-М-специфичности прибегают к их разведению, что вполне приемлемо для рутинного анализа, но не для метода адсорбции - элюции. На активность моноклональных анти-М- и анти-К-антител влияют рН среды и температурные условия постановки реакции. Поликлональные антитела анти-М и анти-N алло- и ксеногенного происхождения менее зависимы от этих условий.
Протеолитические ферменты, применяемые в практике иммуносерологи-ческих лабораторий (папаин, фицин, бромелин, протеаза), разрушают антигены М и N, в связи с чем энзимные пробы при работе с антителами анти-М и анти-N не используют (Hirsch и соавт. [93], Issitt, Anstee [113], Race, Sanger [191], Reid и соавт. [201]).